Caracterizarea microorganismelor utilizate in industria alimentara
- Detalii
- Categorie: Merceologia mărfurilor alimentare
- Accesări: 41,044
În procesele microbiologice industriale microorganismele sunt utilizate pentru următoarele scopuri:
- multiplicarea celulelor în scopul obţinerii de culturi starter sau biomasă;
- supersinteza intra/extracelulară a metaboliţilor primari, metaboliţilor secundari, poliglucide capsulare şi a enzimelor microbiene cu importanţă practică deosebită;
- procese de bioconversie (biotransformarea substraturilor specifice) cu aplicaţii industriale diverse şi depoluarea mediului ambiant.
Utilizarea microorganismelor în bioprocesele industriale este avantajoasă datorită proprietăţilor lor biologice particulare şi anume:
- viteza deosebit de mare a reacţiilor metabolice specifice microorganismelor face ca procesele de biosinteză să aibă loc extrem de rapid comparativ cu procesele similare la plante sau animale;
- se dezvoltă rapid pe diverse medii producând degradări, modificări şi substituiri ale componentelor diferitelor substraturi pe care se dezvoltă şi realizează cu uşurinţă transformări a căror realizare prin procedee chimice ar necesita un număr mare de operaţii laborioase şi o tehnicitate ridicată;
- datorită capacităţii de adaptare la medii cu compoziţii chimice foarte diferite, microorganismele pot utiliza o gamă extinsă de materii prime, uneori lipsite de valoare economică, ceea ce permite utilizarea unor substraturi ieftine pentru obţinerea pe cale biologică a unei game largi de produse utile;
- posibilitatea creşterii randamentului de biosinteză prin metode culturale sau manipulări genetice.
În orice proces microbiologic cultura microbiană utilizată are importanţă primordială în desfăşurarea procesului şi randamentelor obţinute, ea reprezentând cheia succesului sau eşecului proceselor.
Microorganismele utilizate în procesele microbiologice industriale trebuie să posede următoarele calităţi:
- să prezinte tulpini stabile genetic;
- să producă mai multe tipuri de celule (vegetative, spori sau alte unităţi reproductive);
- să nu fie contaminate cu alte microorganisme;
- să crească viguros şi rapid după inoculare în vasele de cultivare utilizate pentru prepararea inoculului înainte de fermentaţia industrială;
- să realizeze bioprocesul dorit într-o perioadă scurtă de timp (preferabil în 3 zile sau mai puţin) şi să aibă capacitatea de a creşte pe medii cu componente ieftine şi uşor de procurat;
- cultura microbiană să poată fi uşor conservată în timp fără a-şi schimba proprietăţile biochimice şi genetice;
- să prezinte stabilitate metabolică şi să asigure desfăşurarea de procese metabolice reproductibile;
- să se preteze la schimbări sub acţiunea agenţilor mutageni sau prin inginerie genetică, în vederea obţinerii de tulpini programate cu randamente sporite de biosinteză şi stabilitate metabolică deosebită.
Microorganisme cele mai utilizate în scopuri industriale aparţin următoarelor grupuri:
- fungi:
- drojdii (levuri);
- mucegaiuri (micromicete, fungi filamentoşi);
- bacterii;
- alge microscopice.
Drojdii (levuri)
Drojdiile reprezintă un grup complex şi eterogen de microorganisme monocelulare de tip eucariot (celule cu nucleu diferenţiat), care se înmulţesc prin înmugurire, ca formă generală de reproducere şi în mod particular prin ascospori formaţi pe cale asexuată şi sexuată.
Denumirea de „drojdie” semnifică, în general, sedimentul care se formează după ce a avut loc fermentaţia alcoolică a musturilor dulci. Denumirea de levuri provine de la verbul „lever” (limba franceză - a ridica) ce sugerează creşterea în volum a aluatului la fabricarea pâinii. Cunoscute din timpuri străvechi pentru activitatea lor fermentativă, drojdiile sunt studiate de Louis Pasteur (1863), studiu continuat de microbiologii: Meyen, Rees şi Hansen care realizează o clasificare a drojdiilor în 1896, completată de Guillermond în 1928. Un studiu taxonomic de referinţă aparţine lui Lodder (1952, 1970) şi mai recent, lui Kreger van Rij (1984).
Importanţă Drojdiilor şi rol
Drojdiile joacă un rol foarte important în industria alimentară, biotehnologie, în natură.
Având drept caracteristică principală capacitatea de a produce fermentarea glucidelor simple cu formare de alcool etilic şi dioxid de carbon, drojdiile fermentative sunt utilizate în biotehnologii alimentare la fabricarea alcoolului, a vinului, berii şi pâinii. Drojdiile au o compoziţie chimică valoroasă şi după cultivare în condiţii de aerare şi prelucrare sunt utilizate ca sursă de proteine în alimentaţia umană, cu denumirea de SCP (single cell protein - proteine din monocelulare) sau în alimentaţia animalelor, deoarece pe lângă 45-55% proteină brută/100 g s.u., aduc în raţie aminoacizi (lizină etc.) şi vitamine ale grupului B (tiamină şi riboflavină).
În microbiologia industrială, din biomasă de drojdie se obţin:
- extracte (plasmolizate, autolizate), folosite ca aditivi alimentari sau pentru îmbogăţirea în substanţe azotate a mediilor de cultură destinate fermentaţiilor;
- vitamine hidrosolubile: B1, B2, PP, ergosterol;
- enzime: β-fructofuranozidaza şi β-galactozidaza;
- prin hibridizări şi inginerie genetică, din mutanţi ai speciei Saccharomyces cerevisiae s-a obţinut interferonul - substanţă cu efect antiviral şi citostatic.
Răspândire drojdiilor în natură
Drojdiile au o largă răspândire în mediul ambiant, fiind întâlnite în toate habitaturile naturale. În sol, celulele de drojdie se întâlnesc în straturile superficiale până la adâncimi de aproximativ 30 cm, în concentraţii de 10 2 - 2·10 5 /g. Din sol, prin acţiunea unor factori (fizici, mecanici, biologici), microorganismele se pot afla temporar în aer şi să se răspândească la distanţe mari; din sol şi aer drojdiile pot ajunge în ape şi unele specii pot fi întâlnite chiar la adâncimi de 4000 m.
În regnul vegetal drojdiile se întâlnesc pe suprafaţa frunzelor, fructelor, florilor, în rădăcini. Răspândirea drojdiilor este favorizată de insecte care odată cu nectarul/sucul preiau şi celulele de drojdie. O parte din drojdii supravieţuiesc iarna în tubul digestiv al insectelor, iar primăvara sunt eliminate. În regnul animal drojdiile sunt prezente în microflora intestinală şi se elimină natural prin produse de dejecţie; în cantităţi mai reduse se întâlnesc în cavitatea bucală şi pe piele.
Caractere morfologice generale
Celula de drojdie are în mod obişnuit formă sferică, ovală sau cilindrică, cu dimensiuni medii de 4-14 µm. Forma şi dimensiunea celulelor este un caracter de gen şi specie şi acestea pot fi influenţate de starea fiziologică şi condiţiile de cultivare. Dintre formele caracteristice unor genuri cu importanţă industrială se menţionează:
- forma ovală (elipsoidală) specifică drojdiilor fermentative ce aparţin genului Saccharomyces;
- forma sferică predomină la drojdii din genul Torulopsis ;
- forma apiculată (de lămâie) la genurile Kloeckera şi Hanseniaspora ;
- forma de sticlă la drojdii din genul Saccharomycodes .
Prin cultivarea celulelor de drojdie pe medii nutritive solide sau solidificate (de exemplu: must de malţ cu agar), în cazul în care la suprafaţă se află o celulă unică, cu caractere morfologice specifice. Coloniile de drojdii au diametrul între 0,2-2 cm au aspect cremos, lucioase sau mate, de culoare în general alb-crem; drojdii din g. Rhodotorula şi Sporobolomyces formează colonii colorate în roşu-portocaliu datorită pigmenţilor carotenoizi sintetizaţi intracelular.
În funcţie de specie, colonia în secţiune poate avea un profil lenticular, semicircular sau triunghiular cu perimetrul circular, umbonat, cu margini ondulate. Caracterele morfocoloniale reprezintă criterii valoroase pentru identificarea drojdiilor.
Structura celulei de drojdie
Ca şi celelalte celule eucariote, drojdiile au structura compusă din învelişuri celulare, citozol, nucleu, mitocondrii, ribozomi, vacuole şi alte organite intracelulare.
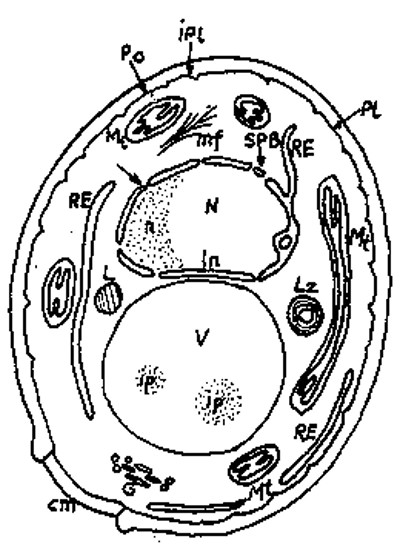
Fig.1. Organizarea internă a celulei de drojdie:
cm - cicatrice mugurală
G - corp Golgi
L - incluziuni lipidice
Lz - lizozomi
Mt - mitocondrie
N - nucleu
n - nucleol
Pc - perete celular
Pl - plasmalemă
RE - reticul endoplasmatic
V - vacuolă
In - înveliş nuclear prevăzut cu pori
Ipl - învaginări plasmalemale
Învelişurile celulare - peretele celular şi plasmalema - sunt structuri ce limitează celula şi determină forma de bază a acesteia şi intervin în toate procesele biologice fundamentale care se desfăşoară la nivel celular.
Peretele celular are o grosime cuprinsă între 150 şi 250 nm şi poate să reprezinte o pondere de 5÷15% din biomasa uscată a drojdiei. Din punct de vedere structural, peretele celular are aspect laminar şi este alcătuit din 2÷3 straturi. Diferenţiat după stadiul de dezvoltare al celulelor, peretele celular se prezintă subţire şi flexibil la celulele tinere, mai gros şi rigid la celulele bătrâne.
Stratul extern prezintă o suprafaţă rugoasă în care se observă cicatricile mugurale, rămase după desprinderea mugurilor de celula mamă. O celulă normală de Saccharomyces cerevisiae poate înmuguri de circa 20 de ori, aşa încât înainte de a-şi pierde viabilitatea aproape întreaga suprafaţă celulară este acoperită cu cicatrici mugurale.
Din punct de vedere chimic, în componenţa stratului extern s-au descoperit manani, glucani şi chitina, care împreună reprezintă peste 90% din substanţa uscată a peretelui celular, alături de prezenţa unor cantităţi mici şi variabile de lipide, enzime, cationi şi apă.
Stratul intern are o suprafaţă ornamentată cu riduri proeminente formate din fibrile constituite din molecule liniare de b-glucan ce formează complexe cu proteinele asigurând rigiditatea sau elasticitatea peretelui celular.
La nivelul peretelui celular sunt localizate şi enzime (invertaza, fosfataza, endoglucanaze, permeaze) implicate în biosinteza compuşilor peretelui celular şi în procese de transfer a substanţelor.
Rolul esenţial al peretelui celular este cel de a asigura forma celulei şi de protecţie faţă de factorii mediului ambiant. În cooperare cu plasmalema, participă la creşterea şi reproducerea celulară, în biosinteză şi catabolism.
De asemenea, peretele celular este implicat în reacţiile de floculare şi aglutinare ale drojdiilor, proprietăţi tehnologice importante pentru drojdiile fermentative.
Prin îndepărtarea peretelui celular prin metode enzimatice, se obţin protoplaştii folosiţi în ingineria genetică pentru obţinerea prin fuziune a hibrizilor cu importanţă practică. Prin amplasarea protoplaştilor pe mediu nutritiv în timp de 8÷12 ore are loc regenerarea peretelui celular.
Plasmalema (membrana citoplasmatică) , reprezintă cel de-al doilea înveliş al celulei de drojdii ce aderă strâns pe toată suprafaţa sa cu peretele celular, cu care cooperează funcţional. La microscopul electronic apare ca un strat lamelar cu o grosime de circa 8÷9 nm, care delimitează protoplastul la exterior. La celulele tinere apare ca un strat omogen, neted, fără nici o învaginare, în timp ce la celulele mature în membrană se evidenţiază învaginări - similare unor şanţuri care îi măresc mult suprafaţa.
Din punct de vedere chimic plasmalema este constituită din lipoproteine, în care lipidele reprezintă 23÷30%, iar proteinele 30÷33% din masa membranei, raportul proteine/lipide variază în funcţie de specie, fenofază şi procedeul de izolare utilizat, între 1,1 şi 1,4. Dintre lipidele prezente în plasmalemă predomină fosfolipidele care împreună cu sterolii şi acizii graşi nesaturaţi au un rol esenţial în procese de permeabilitate.
Lipidele, cu rol de solvent hidrofobic, permit accesul în celulă al unor substanţe utile (aminoacizi nepolari, vitamine). Proteinele plasmatice sunt glicoproteine ce au o mare mobilitate în plasmalemă şi sub formă de molecule sau complexe moleculare îndeplinesc funcţii de recepţie şi transport.
Conform modelului stabilit de Robertson, plasmalema prezintă două straturi lipidice în care moleculele de lipide sunt orientate cu partea hidrofobă spre interior şi cu cea hidrofilă - polară spre periferie şi sunt delimitate în ambele părţi de macromolecule proteice; particule globulare proteice se pot evidenţia şi între straturile lipidice, modificându-şi poziţia în funcţie de starea fiziologică a celulei.
Plasmalema este o structură dinamică, însuşirile sale morfologice şi biochimice ca şi potenţialul său funcţional modificându-se pregnant în funcţie de stadiul ciclului celular şi de influenţa unor factori ai mediului ambiant. În primul rând este o barieră osmotică cu permeabilitate selectivă ce reglează transferul de substanţe nutritive necesare în celulă pentru a fi metabolizate, precum şi eliminarea de cataboliţi, iar prin sistemele enzimatice active la acest nivel, intervine în reglarea procesului de creştere şi înmulţire. Plasmalema interferează cu principalele căi metabolice ale celulei vii.
Citoplasma este constituită din citosol şi din organite citoplasmatice care sunt dispersate în el. Este componenta celulară cu cel mai înalt dinamism, structura sa fizico-chimică şi potenţialul său fiziologic modificându-se foarte rapid, corelându-se permanent cu condiţiile mediului ambiant şi cu necesităţile metabolice ale diferitelor etape de dezvoltare.
Citoplasma reprezintă mai mult de jumătate din volumul total al celulei. Hialoplasma sau matricea citoplasmatică este substanţa fundamentală a citoplasmei prezentă în toate celulele vii, localizată în spaţiul celular în afara compartimentelor delimitate de membranele intracelulare. Ea este formată dintr-o fracţie solubilă (citosol), citoschelet şi o reţea de filamente care interconectează elemente de citoschelet şi organitele celulare în citoplasmă (Dan, V., 1997).
Citosolul este un sistem coloidal cu un conţinut de 70÷85% apă, în care substanţele componente se află sub formă de sol sau gel, formând micele coloidale. Dintre substanţele organice predomină proteinele cu rol structural sau catalitic, lipide cu rol plastic şi glucide cu rol energetic. În compoziţia chimică a citosolului intră şi acizii nuceici (ARN ŞI ADN), nucleosidele şi nucleotidele cu implicaţii majore în metabolismul energetic al celulei ca şi unii alcooli, fenoli, acizi organici, etc. În citosol, în anumite faze ale creşterii celulare, se pot acumula substanţe în exces sub forma unor incluziuni de rezervă şi anume:
- Granule de glicogen, se prezintă sub forma unor corpusculi sferici, cu diametrul de circa 40 nm şi reprezintă principala substanţă de rezervă a celulei care este metabolizată când aceasta este în stare de înfometare. Se pot evidenţia prin suspendarea celulelor de drojdie în soluţie de Lugol, când glicogenul în prezenţa iodului se colorează în brun-roşcat.
- Sferozomii (oleiozomii) reprezintă incluziuni lipidice acumulate în faza staţionară de creştere celulară delimitate de o membrană simplă, ce conţin fosfolipide şi acizi graşi nesaturaţi ce pot fi metabolizaţi în condiţii de malnutriţie, după ce se produce liza enzimatică a membranei.
Citoplasma joacă un rol important în biochimia celulei şi se găseşte în strânsă interdependenţă cu organitele pe care le înconjoară. În acest sens, primul stadiu al respiraţiei - glicoliza - şi fermentaţia alcoolică decurg în citoplasmă, în timp ce următorul stadiu, ciclul ATC se realizează mitocondric.
Organitele celulare
Dintre organitele celulare cu care citosolul interacţionează în mod permanent formând o unitate morfofuncţională, fac parte următoarele: nucleul, mitocondrii, aparatul Golgi, sistemul vacuolar.
Nucleul reprezintă spaţiul genetic în care are loc stocarea, replicarea şi transmiterea informaţiei celulare şi poate fi secvenţializat în următoarele părţi: înveliş, nucleoplasmă, cromonemată, nucleol şi fus intranuclear. Are o formă sferică sau ovală, de obicei cu o poziţie excentrică în citosol şi prezintă o anvelopă nucleară în care s-au evidenţiat pori care asigură legătura între nucleu şi citosol.
Învelişul nuclear conferă individualitate nucleului prin controlul riguros şi medierea schimbărilor nucleocitoplasmatice. Nucleoplasma reprezintă mediul în care sunt înglobate cromonemata, nucleolul şi fusul intranuclear. Nucleolul constă dintr-un miez denumit core, alcătuit din histone în formă de disc pe care sunt înfăşurate segmente de ADN, formând aranjamente helicoidale sau solenoide.
În nucleol sunt sintetizate proteine ribozomale şi ARN ribozomal care apoi trec prin porii nucleari, în citosol. În funcţie de caracterele genetice ale speciei de drojdie, numărul de cromozomi nu depăşeşte valori de 8÷18, Saccharomyces cerevisiae are 17 cromozomi (Konovalov, 1980).
Mitocondriile sunt organitele de bază sau structurile subcelulare în care se sintetizează substanţele care înmagazinează energia chimică a celulei, de aceea, nu întâmplător sunt supranumite şi „uzine energetice celulare”. Pot ocupa până la 25% din volumul citosolului, în număr variabil, de 10÷50/celulă, într-o continuă metamorfoză, încât în unele faze fiziologice pot forma o mitocondrie unică, ramificată denumită condriom.
Sunt organite puternic hidratate, apa reprezentând până la 70% din masa mitocondriei, ceea ce reflectă un potenţial fiziologic particular. Substanţa uscată este constituită în principal din proteine şi lipide (peste 90%), acizi nucleici, nucleotide, ioni minerali. Proporţia diferitelor componente variază considerabil în funcţie de starea fiziologică a celulei şi de influenţa unor factori de mediu. Conform ultimelor date ştiinţifice, proteinele se află repartizate în proporţie de 5% în membrana externă, 20% în membrana internă şi 75% în condriopasmă.
Mitocondriile posedă sisteme de transport specifice, diversificate, care realizează transportul unor metaboliţi esenţiali prin membranele mitocondriale ce separă condrioplasma de citosol. În mitocondrii există enzime ce realizează procese de oxidare ale compuşilor organici, enzimele lanţului respirator şi numeroase enzime oxidative, enzime ce asigură fosforilarea.
Reticulul endoplasmatic face legătura între nucleu şi vacuom şi reprezintă o reţea de vezicule, caracterizate printr-o mare plasticitate morfologică. Reticulul endoplasmatic este sediul unor complexe enzimatice (citocromoxidaza, NAD + , enzime ale lanţului transportor de electroni) şi are rol în transportul substanţelor nutritive, în declanşarea procesului de înmugurire, în biogeneza sferozomilor, vacuolelor, corpilor Golgi; participă, de asemenea, la expansiunea învelişului nuclear şi a plasmalemei în diferite etape de dezvoltare ale celulei.
Aparatul Golgi este un sistem de endomembrane, alcătuit din unităţi funcţionale - corpi Golgi sau dictiozomi, care face legătura între reticulul endoplasmatic şi plasmalemă, între reticulul endoplasmatic şi alte organite celulare. Aparatul Golgi este aparatul de sortare şi dirijare a proteinelor şi componentelor membranare spre locul de destinaţie; au rol în expansiunea peretelui celular (Dan, V., 1999).
Ribozomii denumiţi şi granule Palade după numele cercetătorului de origine română care i-a pus în evidenţă în anul 1953, sunt particule nucleoproteice implicate în sinteza proteinelor celulare răspândiţi în citosol - citoribozomi, liberi sau în asociaţii de 5÷6 polizomi. Au în structură ARN - ribozomal şi proteine (în cazul ribozomilor de Sachharomyces cerevisiae valoarea raportului ARN/proteine variază între 1,04 şi 1,12. ARN ribozomal este implicat în procese de transcripţie a informaţiei genetice pentru biosinteza proteinelor/enzimelor, necesare celulei.
Lizozomii sunt structuri veziculare bogate în enzime: proteaze, fosfataze, lipaze (peste 40 de tipuri de enzime), active la pH = 5 cu rol în digestia unor compuşi ai celulei vii, care nu mai funcţionează eficient; în exteriorul lizozomului, în citosol, enzimele nu sunt active deoarece pH-ul este de 7,3. Enzimele lizozomale sunt în mod normal inactive, latenţa lor este dependentă de integritatea membranei lizozomale, care acţionează ca o barieră ce se interpune între enzimă şi substrat. Când sub acţiunea unor factori, de exemplu în starea de înfometare, în absenţa apei care să asigure transportul în exteriorul celulei a cataboliţilor formaţi, pH-ul în citosol scade, enzimele din lizozomi sunt activate şi celula moare prin autoliză.
Peroxizomii sunt structuri sferice sau ovale înconjurate de o membrană simplă şi cu o matrice fin granulată pe care sunt localizate oxidaze, cu rol în adaptarea celulei de drojdie la condiţii aerobe şi intervin în degradarea acizilor graşi, a aminoacizilor, a apei oxigenate şi a altor substanţe (de ex. a metanolului).
Caractere fiziologice generale ale drojdiilor
O proprietate importantă a unor drojdii cu utilizări în industria alimentară este aceea de a fermenta în condiţii de anaerobioză glucide (hexoze, diglucie, triglucide) cu formare de alcool etilic şi dioxid de carbon şi produse secundare care dau aroma caracteristică produselor fermentate:
C 6 H 12 O 6 -----→ 2CH 3 -CH 2 -OH + 2CO 2 + 117kj
În condiţii de aerobioză, drojdiile asimilează glucidele transformându-le prin respiraţie la CO 2 şi H 2 O, iar energia eliberată favorizează creşterea şi înmulţirea celulelor:
C 6 H 12 O 6 ———→ 6CO 2 + 6H 2 O + 2840 kj
Celulele de drojdie reacţionează activ la presiunea osmotică dată de concentraţia substanţelor dizolvate în mediul în care se află suspendate. Dacă mediul este hipotonic, cu o concentraţie a substanţelor dizolvate mai mică decât concentraţia intracelulară, apa va pătrunde în celula care îşi măreşte volumul şi dacă se prelungeşte această stare de turgescenţă, celula suferă deteriorări fizice ireversibile. În mediu hipertonic, când concentraţia mediului este superioară concentraţiei intracelulare, apa din celulă difuzează în exterior pentru a asigura izotonia, iar celula trece în starea de plasmoliză (Dan, V., 1999).
Celulele de drojdie, în funcţie de condiţiile de cultivare si vârstă pot prezenta activitate metabolică diferenţiată concretizată prin trei stări în care se pot afla celulele si anume:
- starea de metabioză (activă), în care celulele au activitate metabolică maximă. În condiţii favorabile de cultură, celulele cresc, se reproduc şi acumulează intracelular substanţe de rezervă glicogen şi trehaloză;
- starea de anabioză (latentă), celulele îşi menţin caracteristicile vitale, dar nu se pot înmulţi. Această stare apare când substanţele nutritive ale mediului sunt epuizate, iar celulele consumă din substanţele de rezervă intracelulare;
- starea de autoliză (moartea fiziologică). Se produce o solubilizare a compuşilor intracelulari sub acţiunea enzimelor proprii celulei.
Reproducerea drojdiilor
Forma generală de reproducere a drojdiilor este înmugurirea vegetativă (calea asexuată) care are la bază un proces simplificat de mitoză, când din celula mamă se formează noua celulă, identică din punct de vedere genetic.
Unele drojdii au capacitatea de a se înmulţi prin sporulare, ce poate avea loc pe cale asexuată (prin procesul de meioză) şi pe cale sexuată (prin procese de copulare). Sporularea este condiţionată genetic are loc în anumite condiţii de mediu.
Reproducerea prin înmugurire. Înmugurirea are loc când drojdiile se află în condiţii optime de mediu (nutrienţi, pH, temperatură). O condiţie necesară pentru înmugurire este aerarea corespunzătoare a mediului, deoarece în prezenţa oxigenului din aer are loc asimilarea eficientă a nutrienţilor cu recuperarea energiei potenţiale a acestora, energie folosită de celulă în creştere pentru procese de biosinteză, consumatoare de energie.
În aceste condiţii, în prima etapă celula de drojdie creşte în dimensiuni în urma creşterii coordonate a compuşilor intracelulari; creşterea în volum fiind mai rapidă decât a suprafeţei învelişurilor celulare, are loc o înmuiere enzimatică a peretelui şi apare o protuberanţă - mugurele. Între celula parentală şi mugure se perfectează un canal - diverticulum prin care se transferă în celula nou formată material nuclear şi citoplasmatic (cariokineză şi citokineză).
Când celula fiică va conţine toate componentele şi organitele necesare unei vieţi independente, la nivelul canalului se formează un perete inelar cu o concentraţie ridicată în chitină, ce se dezvoltă centripet până are loc obturarea şi separarea (Dan, V., 1999).
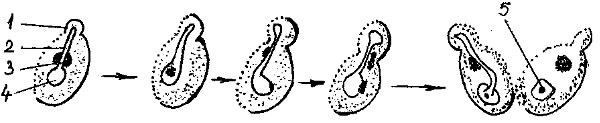
Fig. 2. Înmugurirea la drojdii:
-
mugure; 2- canal de legătură; 3- nucleu; 4- vacuolă adiacentă nucleului; 5- nucleol.
Celula fiică, cu activitate metabolică independentă, poate rămâne ataşată de celula mamă formând asociaţii sau pseudomicelii, sau se separă de celula parentală, iar la locul de desprindere rămâne vizibilă cicatricea mugurală. În condiţii optime de viaţă, o celulă de drojdie poate forma 9÷42 noi celule apoi celula moare din punct de vedere fiziologic.
Reproducerea prin sporulare. Sporularea are la bază procesul de meioză, prin care numărul de cromozomi ai celulei parentale diploide se reduce la jumătate, iar celulele formate primind un singur rând de cromozomi sunt haploide.
Sporularea este iniţiată după circa 10 ore de expunere pe mediul de sporulare şi este mai intensă după 24÷48 ore. În timp, celula generatoare de spori denumită şi asc se poate rupe sau să se solubilizeze prin autoliză şi ascosporii liberi, în condiţii favorabile germinează şi formează prin reproducere vegetativă clone haploide. Ascosporii haploizi pot fi de tip “a” sau “α” în funcţie de natura feronomilor specifici; în condiţii favorabile, celulele ajunse la maturitate fiziologică denumite şi gameţi pot să fuzioneze printr-un proces de conjugare pentru a forma din nou celule diploide. Dacă are loc conjugarea între celule de acelaşi tip “a” sau “α” celulele diploide sunt viabile, dar ascosporii formaţi îşi pierd această calitate.
Sub formă de ascospori drojdiile rezistă timp îndelungat în sol, în condiţii de uscăciune, în schimb termorezistenţa lor este apropiată cu cea a celulelor vegetative (inactivare la 75÷85 0 C).
Meioza este importantă din punct de vedere practic pentru că se pot realiza copulări dirijate cu obţinerea unor hibrizi ce prezintă proprietăţi superioare preluând de la celulele parentale caractere deosebite (Dan, V., 1999).
Clasificarea generală a drojdiilor
Potrivit Codului Internaţional de Nomenclatură Botanică tulpinile de drojdii sunt grupate în specii, speciile în genuri, iar genurile în subfamilii şi familii.
O clasificare de referinţă este cea propusă de Lodder şi Kreger van Rij (1952) care clasifică drojdiile în 38 genuri şi 349 de specii de drojdii, iar clasificarea cea mai recentă şi cuprinzătoare aparţine lui Kreger van Rij (1984) (60 genuri cu aproximativ 500 de specii).
Din această clasificare se vor menţiona genuri şi specii cu implicaţii în industria alimentară, în calitatea lor de agenţi ai fermentaţiei alcoolice sau de alterare a produselor alimentare.
Genul Saccharomyces cuprinde 45 de specii cu activitate predominant fermentativă. Se înmulţesc prin înmugurire şi sporulare, producând 1-4 ascospori. Dintre speciile reprezentative ale genului:
Saccharomyces cerevisiae este o drojdie de fermentaţie superioară folosită la fabricarea alcoolului şi la obţinerea drojdiei de panificaţie. Din biomasa de celule obţinută în mediul nutritiv şi în condiţii de aerare, prin procedee biotehnologice se pot obţine enzime (invertază), vitamine din grupul B, interferon ş.a.
Saccharomyces carlsbergensis este o drojdie de fermentaţie inferioară, se utilizează industrial la fabricarea berii, iar din drojdia reziduală rezultată după fermentare se pot obţine extracte, substanţe de aromă.
Genul Brettanomyces cuprinde drojdii de formă oval cilindrică, în aerobioză pot să producă prin fermentarea lentă a glucozei, alcool etilic 11-12% alcool, acid acetic, citric, lactic, succinic. Dau alterări ale berii, vinului (tulburare, iz de şoarece) cu formare de esteri şi substanţe cu gust amar, a băuturilor nealcoolice, a murăturilor.
Genul Candida este un gen bogat în specii (81), eterogen din punct de vedere morfologic şi fiziologic şi care a suferit în timp multe modificări taxonomice. Candida mycoderma denumită şi floarea vinului se dezvoltă în prezenţa aerului la suprafaţa lichidelor slab alcoolice formând un voal caracteristic; prin oxidarea alcoolului la dioxid de carbon şi apă are loc deprecierea vinului, berii.
Alte specii Candida utilis, Candida robusta, Candida tropicalis, Candida lypolitica etc. se pot cultiva pe medii obţinute prin prelucrarea unor deşeuri ale industriei alimentare, a lemnului şi celulozei, obţinându-se o biomasă, cu un conţinut de 45-55% proteină, folosită în furajarea animalelor. Candida kefyri este utilă la fabricarea chefirului, Candida albicans , drojdie facultativ patogenă nu se înmulţeşte în alimente.
Genul Debaryomyces cuprinde drojdii de formă oval cilindrică, poate produce mucus la suprafaţa batoanelor de salam, se dezvoltă la suprafaţa brânzeturilor, iaurtului şi dau alterări ale sucurilor concentrate de portocale.
Genul Hanseniaspora include drojdii cu proprietăţi fermentative reduse (5 0 alcool), cu formă apiculată şi sunt răspândite în microbiota fructelor citrice, a smochinelor; intervin în fermentarea naturală a boabelor de cacao.
Hanseniaspora apiculata poate reprezenta până la 90% din microbiota mustului şi îşi reduce viteza de fermentare la acumularea de 5-7% alcool. Se poate dezvolta în vin fiind responsabilă pentru formarea de acizi volatili, esteri care dau un gust amar, străin vinului. Prin contaminarea vinului folosit la obţinerea şampaniei la sticle, Hanseniaspora apiculata poate forma un sediment aderent la sticlă şi care se îndepărtează greu la degorjare.
Genul Pichia conţine 35 de specii. Au o slabă activitate fermentativă, sunt drojdii peliculare; prin dezvoltare şi formare de voal produc deprecierea vinului, a berii. Se pot izola de pe suprafaţa peştilor şi a creveţilor, din saramura măslinelor conservate şi pot produce alterări ale produselor vegetale murate.
Genul Rhodotorula include celule cu formă oval-cilindrică, sunt drojdii oxidative şi pot sintetiza pigmenţi carotenoizi care imprimă coloniei culoarea roşu-cărămiziu. Rhodotorula glutinis şi Rhodotorula mucilaginosa , predominante pe alimente: pui, peşte, creveţi, pe suprafaţa untului, sunt specii psihrotrofe.
Genul Saccharomycodes cu specia Saccharomycodes ludwigii prezintă forme apiculate sau oval alungite, poate fi agent de alterare a musturilor şi sucurilor de fructe şi dau tulburarea cidrului, a vinurilor semidulci.
Genul Saccharomycopsis cu specia importantă Saccharomycopsis fibuligera prezintă celule ovale, fermentează lent glucoza, zaharoza, maltoza, asimilează amidon, alcool etilic, acid lactic etc.; unele tulpini sunt folosite pentru producerea de glucoamilază sau pentru obţinerea de drojdii furajere.
Genul Schizosaccharomyces cuprinde drojdii de formă cilindrică, ovală. Schizosaccharomyces pombé este agent al fermentării sucului de trestie de zahăr şi folosită pentru obţinerea romului şi a băuturii arak.
Genul Torulopsis prezintă celule sferice, ovale sau cilindrice. Specii ale genului întâlnite în mustul de struguri, au putere alcooligenă redusă. Sunt osmotolerante, psihrofile, sulfitorezistente. Pot produce alterări ale laptelui concentrat, a siropurilor, a sucurilor.
Genul Zygosaccharomyces cuprinde drojdii cu celule ovale, cu proprietăţi fermentative, osmotolerante. Pot produce fermentarea mierii, a siropurilor concentrate de zahăr cu formare de alcool etilic, acid acetic, dioxid de carbon.
Mucegaiuri (fungi filamentoşi, micromicete)
Mucegaiurile sunt microorganisme de tip eucariot, monocelulare sau pluricelulare, diferenţiate din punct de vedere morfologic şi care se reproduc prin spori formaţi numai pe cale asexuată sau pe cale mixtă (asexuată şi sexuată).
Răspândire şi rol
Mucegaiurile sunt foarte răspândite în natură, habitatul preferat şi cel mai populat este solul, în special în stratul superficial al solului care le asigură condiţii de creştere şi supravieţuire. Prin activitatea lor de degradare a materiei organice vii, mucegaiurile participă la transformarea unor compuşi organici (celuloza, hemiceluloze, substanţe pectice, amidon, lipide) la compuşi mai simpli şi sunt consideraţi agenţi de putrezire.
Mucegaiurile participă astfel la circuitul carbonului în natură şi îmbogăţesc solul în substanţe cu molecule mici care pot fi folosite de alte microorganisme sau de către plante. Din sol, prin intermediul factorilor naturali, sporii de mucegai sunt antrenaţi pe calea aerului la distanţe foarte mari, ceea ce asigură răspândirea nelimitată de graniţe geografice. În aer, mucegaiurile sub formă de spori sau hife vegetative pot supravieţui un timp îndelungat, iar în absenţa curenţilor de aer se depun cu o viteză ce atinge valori de 3 cm/sec.
În apă prezenţa mucegaiurilor este ocazională, apa fiind un mediu prin care se poate face răspândirea sporilor. Creşterea mucegaiurilor în apă este dependentă de conţinutul acestora în compuşi organici şi poate avea loc numai în condiţii de aerare.
Sporii de mucegai se întâlnesc frecvent la suprafaţa plantelor, în tractul digestiv, în special la ierbivore.
În industria alimentară activitatea de biodeteriorare este nedorită deoarece fungii cauzează pierderi prin mucegăirea seminţelor alimentelor. Ca efect secundar este formarea de micotoxine, de către unele mucegaiuri, încât alimentele devin inutilizabile.În afara mucegaiurilor saprofite - agenţi ai putrezirii, se întâlnesc mucegaiuri patogene care pot parazita: plante, animale, peşti şi insecte.
Mucegaiurile sunt frecvent întâlnite în microbiota plantelor, pe suprafaţa fructelor şi legumelor. Dacă produc îmbolnăviri la plante poartă denumirea de mucegaiuri fitopatogene şi sunt responsabile pentru aproximativ 70% din totalul îmbolnăvirilor întâlnite la cereale şi legume. Dintre mucegaiurile fitopatogene se pot enumera cele care produc boli ca mălura, rugina, tăciunele etc. ale plantelor industriale.
La om şi animale mucegaiurile patogene produc un număr mai redus de îmbolnăviri, se dezvoltă pe piele, unghii, păr. Un număr mic de mucegaiuri pot produce îmbolnăviri interne atunci când sporii sunt inhalaţi (pe cale respiratorie) dând micoze (de exemplu, Aspergillus fumigatus produce aspergillom pulmonar).
Rolul mucegaiurilor în industrie
În afară de rolul important al mucegaiurilor în natură, în industria alimentară, culturi fungice selecţionate se pot folosi la fabricarea brânzeturilor - tip Roqueforti, Camemberti sau la maturarea salamurilor crude.
Cu ajutorul mucegaiurilor pe cale biotehnologică se pot obţine compuşi deosebit de valoroşi:
- antibiotice: peniciline cu Penicillium chrysogenum , cephalosporine ( Cephalosporium ), griseofulvine ( Penicillium griseofulvum ) acidul fusidic cu Fusidium coccinenum şi antibiotice active faţă de bacterii Gram pozitive cu Mucor ramannianus ;
- acizi organici (citric, lactic, gluconic, kojic, malic, fumaric);
- vitamine (B 2 , ergosterol - provitamina D 2 );
- enzime (amilaze, proteaze, lipaze, celulaze etc.).
Mucegaiurile se mai pot folosi pentru îmbogăţirea în proteine a făinurilor vegetale şi ca agenţi de depoluare a apelor reziduale.
În tehnologiile moderne se obţin SCP cu fungi din care: Fusarium graminearum în Marea Britanie şi specii de Paecilomyces în procesul Pekilo în Finlanda unde sunt cultivate pe leşii sulfitice şi deşeuri din industria hârtiei.
Caractere morfologice
Mucegaiurile se răspândesc în natură sub formă de spori rezistenţi la uscăciune, formă în care se menţin în stare viabilă ani de zile. Dacă un astfel de spor ajunge pe suprafaţa unui mediu favorabil pentru creştere, cu o cantitate suficientă de apă liberă care să-i permită absorbţia substanţelor nutritive, în primul stadiu care poate să dureze 3-4 ore, are loc absorbţia apei şi activizarea sistemelor enzimatice, apoi are loc germinarea celulei sporale şi formarea tuburilor vegetative numite hife sau thal. Hifele cresc numai prin vârf şi deci au o creştere apicală, după care se ramifică.
Hifele se extind pe suprafaţa mediului, se diversifică şi îndeplinesc anumite funcţii specializate. Hifele de extindere se pot dezvolta şi în profunzimea mediului realizând absorbţia nutrienţilor şi au rol în susţinere; hifele de răspândire se pot dezvolta de-a lungul mediului sau aerian. La un anumit grad de dezvoltare a acestor hife vegetative se formează hifele reproducătoare, generatoare de spori, diferenţiate în funcţie de gen şi specie. Totalitatea hifelor vegetative şi reproducătoare alcătuieşte miceliul.
Dezvoltarea mucegaiurilor are loc destul de rapid în condiţii favorabile, astfel în interval de 2-3 zile pe mediu nutritiv se formează colonii vizibile diferenţiate.
În cazul mucegaiurilor inferioare caracterizate prin miceliu aseptat, coloniile se dezvoltă rapid, sunt extinse, cu tendinţa de a ocupa tot spaţiul disponibil, au aspect pâslos şi culori alb, bej, cenuşiu, brun.
Mucegaiurile superioare caracterizate prin miceliu septat formează colonii cu o creştere radială limitată şi culoare ce diferă de la alb la galben, brun, verde, portocaliu, albastru, cu diferite nuanţe specifice în funcţie de gen şi specie.
Când sporii de mucegai ajung la suprafaţa unui mediu nutritiv lichid, prin întrepătrunderea hifelor vegetative se formează o peliculă denumită dermă, la început netedă, în timp, prin creşterea suprafeţei derma se cutează şi la suprafaţă se dezvoltă hifele reproducătoare, se produce sporularea şi colorarea specifică. Prin introducerea sporilor de mucegai în mediu lichid şi cultivarea pe agitator rotativ, ca rezultat al agitării, mucegaiurile formează prin creştere vegetativă, sfere vizibile, întotdeauna de culoare albă.
Structura mucegaiurilor
Mucegaiurile au la bază celula de tip eucariot ce include toate organitele descrise la celula de drojdie. Spre deosebire de drojdii, peretele celular este mai gros şi în afară de α şi β-glucani se găseşte în cantitate mare celuloza. Celula poate conţine 1-2 nuclei cu câte 2-4 cromozomi fiecare. În funcţie de caracterele genetice pot fi:
- mucegaiuri inferioare - sunt monocelulare şi se dezvoltă sub forma unei celule gigantice ce prezintă mai multe ramuri. În aceste celule migrează liber nucleii citoplasmatici şi organitele intracelulare. În cazul în care un spor ajunge pe un mediu nutritiv, atunci prin creştere are loc germinarea sporului care constă într-o creştere în volum a acestuia producându-se hife vegetative. Prima dată se formează hife de extindere care se dezvoltă în paralel de-a lungul mediului nutritiv. Se formează apoi hife submerse ce se dezvoltă în profunzimea mediului realizând absorbţia nutrienţilor şi au rol de susţinere. Totalitatea acestor hife de extindere şi submerse formează miceliul vegetativ. Odată cu creşterea şi dezvoltarea miceliului vegetativ încep să apară hife aeriene care sunt şi reproducătoare;
- mucegaiuri superioare sunt pluricelulare şi au miceliu septat, aceasta pentru că la anumite distanţe apar pereţi despărţitori (septum) prevăzut cu un por central prin care se poate face transfer citoplasmatic.
Caractere fiziologice generale
Mucegaiurile sunt microorganisme uşor adaptabile, deoarece au capacitatea de a forma enzime induse în funcţie de natura substratului pe care se află, încât produc degradarea atât a produselor alimentare cât şi a fibrelor textile, a cauciucului, betonului etc.
Mucegaiurile sub formă de hife sau spori sunt foarte rezistente la uscăciune şi se menţin în stare latentă de viaţă un timp îndelungat.
Mucegaiurile sunt puţin pretenţioase la cantitatea de apă liberă prezentă în produs, de aceea ele pot produce mucegăirea produselor care se conservă prin uscare numai când acestea încep să absoarbă apă din mediul ambiant.
În raport cu oxigenul, mucegaiurile sunt microorganisme aerobe deci necesită pentru creştere prezenţa oxigenului din aer sau a oxigenului dizolvat în mediul lichid.
Mucegaiurile se pot dezvolta în limite largi de pH (1,5÷9) cu o valoare optimă în domeniul acid (pH = 5,5÷6).
Mucegaiurile sunt microorganisme mezofile cu temperaturi optime de creştere la 25ºC, un număr restrâns sunt termofile - cele patogene au temperatura optimă la 37ºC, iar altele sunt adaptate la temperaturi scăzute (0÷3ºC). Rezistenţa termică a mucegaiurilor sub formă de hife sau spori este mică, majoritatea sunt inactivate la temperaturi de 80ºC, cei mai rezistenţi spori sunt distruşi la 88ºC în 10 minute.
Reproducerea mucegaiurilor
Mucegaiurile se înmulţesc pe cale vegetativă şi prin sporulare.
Reproducerea vegetativă se realizează prin intermediul fragmentelor de hife rupte sub acţiunea unor factori mecanici atunci când acestea conţin cel puţin o celulă. Fragmentele hifale chiar când, de exemplu în cazul mucegaiurilor superioare conţin mai multe celule, vor forma o singură colonie. Din acest motiv la determinarea numărului de mucegaiuri din diferite produse exprimarea se face în unităţi formatoare de colonii (UFC).
Fungii filamentoşi pot să crească în dimensiuni fără să modifice raportul între volumul de citoplasmă şi suprafaţa hifelor, astfel încât schimbul de substanţe între miceliu şi mediu implică transport numai pe distanţe scurte.
Se cunosc 3 mecanisme prin care are loc reglarea creşterii miceliului şi anume: prin reglarea extinderii hifelor, prin iniţierea de ramificaţii şi prin distribuirea spaţială a hifelor. Timpul de dublare a miceliului ca şi intervalul între cicluri succesive de formare a septumului depind de specie şi condiţii de cultură şi poate dura aproximativ 2 ore ( Aspergillus nidulans ). Se apreciază că, pentru mitoza completă a nucleilor la mucegaiuri sunt suficiente 10 minute, iar intervalul între mitoză şi apariţia septumurilor este de 20÷40 minute.
Reproducerea prin sporulare . Este forma cea mai răspândită la mucegaiuri şi poate avea loc pe cale asexuată sau pe cale mixtă.
Clasificarea generală a mucegaiurilor
Mucegaiurile reprezintă un grup taxonomic complex, motiv pentru care clasificarea acestora este în continuă transformarea. Numărul posibil de specii ce ar putea exista în natură este apreciat la aproximativ 250000. Mucegaiurile de interes alimentar sunt grupate în 20 de genuri şi aproximativ 1000 de specii. Clasificarea are la bază anumite criterii morfologice, structură, caractere coloniale, pigmentogeneză, integrate cu date fiziologice şi genetice.
Mucegaiurile fac parte din diviziunea eumycota cu următoarele subdiviziuni:
Mastigomycotina
Mastigomycotina - cuprinde mucegaiuri inferioare cu miceliu aseptat care se reproduc prin oospori, pe cale sexuată, cu următoarele genuri: Genul Peronospora - cu specii fitopatogene, produc mana viţei de vie. Genul Phytium - include agenţi ce produc putrezirea plantelor de grâu tinere. Genul Phytophtora - cu specia Phytophtora infestans , agentul producător al manei la cartofi.
Zygomycotina
Zygomycotina - cuprinde mucegaiuri inferioare cu miceliu aseptat care se reproduc prin zigospori pe cale sexuată şi prin sporangiospori şi chlamidospori pe cale asexuată.
Genul Mucor (88 specii) - se caracterizează prin formarea de sporangiospori în stilosporange, la maturitate prin ruperea membranei şi eliberarea sporilor rămâne la baza columelei, un collar. În funcţie de specie, columela poate avea dimensiuni şi forme diferite. Dintre speciile mai importante ale genului: Mucor mucedo , denumit şi mucegaiul alb al pâinii, Mucor racemosus , agent de putrezire a fructelor şi legumelor, Mucor pussillus şi Mucor miehei , specii selecţionate pentru obţinerea de proteaze cu acţiune similară cu cea a cheagului animal.
Genul Rhizopus (11 specii) se caracterizează prin stilosporange de dimensiuni mari, cu columelă semisferică, fără collar după ruperea membranei sporangelui. Sporangiosporii se dezvoltă în mănunchi dintr-un punct în care se dezvoltă rhizoizi - hife de susţinere cu rol absorbant. Extinderea coloniei are loc rapid ca urmare a formării unor lăstari micelieni denumiţi stoloni. Specia cea mai răspândită pe toate produsele alimentare este Rhizopus stolonifer , agent de mucegăire a fructelor şi legumelor. Tulpini selecţionate pot fi folosite pentru obţinerea pe cale fermentativă a acidului fumaric.
Genul Thamnidium - se caracterizează prin formarea de sporangiofori terminaţi cu un stilosporange mare sub care se dezvoltă sporangiofori scurţi purtători de sporangioli cu un număr mic de sporangiospori. Thamnidium elegans produce mucegăirea produselor conservate prin refrigerare.
Genul Absidia prezintă sporangi mici cu columelă de formă conică. Unele specii sunt termofile şi produc îmbolnăviri la animale şi om. Poate produce mucegăirea porumbului şi elaborează toxine.
Ascomycotina
Ascomycotina cuprinde mucegaiuri superioare cu miceliu septat care se reproduc pe cale asexuată şi sexuat prin ascospori.
Genul Byssochlamys produce asci cu 8 ascospori termorezistenţi şi produc alterarea alimentelor conservate cu acizi. Byssochlamys fulva şi Byssochlamys nivea produc alterarea conservelor de fructe.
Genul Monascus cu specia Monascus ruber este folosit pentru obţinerea de coloranţi roşii de uz alimentar.
Basidiomycotina
Basidiomycotina cuprinde mucegaiuri superioare cu ciclu de viaţă mai evoluat şi care se reproduc pe cale sexuată prin bazidiospori.
Genul Puccinia cuprinde specii fitopatogene agenţi ai ruginii cerealelor.
Genul Ustilago produc la grâu şi porumb boala denumită popular tăciune.
Deuteromycotina
Deuteromycotina cuprinde numeroase genuri şi specii de mucegaiuri superioare care se reproduc prin conidiospori şi la care nu există cale sexuată de sporulare sau aceasta nu a fost încă evidenţiată.
Genul Aspergillus (132 specii) cuprinde numeroase specii cu importanţă biotehnologică. Se caracterizează prin formarea de conidiofori drepţi, neramificaţi care poartă capul conidial alcătuit dintr-un suport anatomic denumit veziculă pe care se dezvoltă celulele conidiogene - respectiv fialide, generatoare de lanţuri lungi de fialospori.
Specia Aspergillus niger formează colonii radiale de culoare brun-negru, prezintă două rânduri de fialide pe toată suprafaţa veziculei, capul conidial este sferic. Numeroase tulpini selecţionate sunt folosite pentru obţinerea de enzime: amilaze, proteaze, glucozoxidaze, invertaze, enzime pectolitice sau pentru obţinerea acizilor organici: acid citric, acid lactic, gluconic.
Specia Aspergillus oryzae formează colonii de culoare bej-oranj cu conidiofori drepţi şi cap conidial sferic cu un singur rând de fialide. Este numit pe drept cuvânt „arsenalul enzimelor” deoarece se cunosc peste 200 de enzime elaborate de mucegai şi obţinute în stare purificată. Pentru obţinerea de amilaze mai pot fi folosite speciile Aspergillus awamori, Aspergillus phoenicis, Aspegillus usamii, Aspergillus cinnamommeus .
Aspergillus flavus formează colonii de culoare alb-gălbui la maturitate galben-verzui spre brun cu revers colorat în galben brun. Este răspândit în sol, pe produse vegetale şi are capacitatea de a produce aflatoxine, micotoxine cu efect cancerigen.
Genul Penicillium (453 specii) se caracterizează prin formarea unui aparat reproducător ramificat alcătuit din ram, metule, fialide şi fialospori cu diferenţieri morfologice în funcţie de specie. În cadrul genului, specii selecţionate sunt folosite la obţinerea brânzeturilor cu pastă albastră - Penicillium roqueforti , a brânzeturilor cu pastă moale - Penicillium camemberti , la maturarea salamurilor crude uscate - Penicillium nalgiovense . Pentru obţinerea de antibiotice din grupa penicilinelor se fosesc tulpini de Penicillium notatum şi Penicillium chrysogenum . Numeroase specii sunt agenţi de putrezire şi pot produce micotoxine: Penicillium expansum , Penicillium islandicum, Penicillium citrinum etc.
Genul Botrytis formează colonii extinse, pâsloase, de culoare cenuşie. Botrytis cinerea este denumit mucegaiul cenuşiu şi poate produce putrezirea vulgară sau nobilă a strugurilor. Specii fitopatogene dau boli la floarea soarelui şi alterări în depozit ale fructelor şi legumelor.
Genul Fusarium include specii saprofite răspândite în sol şi specii patogene parazite ale plantelor superioare. Dau putrezirea brună a fructelor citrice, putrezirea umedă a smochinelor, mucegăirea cerealelor (orz, grâu) cu producerea de micotoxine - trichothecene: Fusarium graminearum , Fusarium moniliforme , Fusarium nivali etc.
Genul Cladosporium formează colonii cu aspect catifelat de culoare brun-oliv cu revers colorat în bleumarin-negru. Se reproduc prin blastospori în formă de lămâie. Este prezent în microbiota cerealelor proaspăt recoltate. Este agent al putrezirii negre a strugurilor şi pepenilor galbeni.
Genul Alternaria formează colonii pufoase cu miceliu septat şi conidii mari cu septumuri longitudinale şi transversale. Dau putrezirea brună a fructelor. Este considerat mucegai de câmp şi este prezent pe suprafaţa seminţelor proaspăt recoltate fiind folosit ca indice de prospeţime a cerealelor.
Genul Geotrichum formează colonii extinse, catifelate de culoare albă. Produce miceliu septat din care se separă arthrospori ce au tendinţa de aranjare în zig-zag. Geotrichum candidum este întâlnit în industria laptelui şi la fabricarea pastei de tomate, drept contaminant al utilajelor.
Genul Trichoderma formează colonii extinse pufoase sau pulverulente de culoare gălbui spre verde. Trichoderma reesei produce activ celulaze şi un antibiotic gliotoxina cu efect fungistatic faţă de mucegaiuri care produc putrezirea lemnului.
Genul Trichothecium formează colonii cu aspect pufos de culoare roz-portocaliu. Dintre cele 4 specii ale genului Trichothecium roseum este cel mai întâlnit pe reziduuri vegetale, ca agent al putrezirii fructelor. Este întâlnit pe suprafaţa boabelor de cereale: grâu, orz, porumb şi poate produce mucegăirea pâinii.
Bacterii
Bacteriile sunt microorganisme monocelulare de tip procariot cu un cromozom unic, cu dimensiuni medii între 0,5-0,8 μm, care se înmulţesc asexuat prin sciziune binară, izomoforfă.
Bacteriile sunt microorganisme cu o largă răspândire în natură, ca rezultat al adaptării lor în cursul procesului de evoluţie. Rezervorul natural al bacteriilor este solul unde concentraţia de celule poate ajunge la valori de 10 7 -10 9 g -1 atât în straturile superficiale (bacterii aerobe), cât şi în straturile de profunzime (bacterii anaerobe). Din sol, bacteriile s-au adaptat să trăiască în ape, unde concentraţia de celule poate fi între 10/cm 3 în apa de izvor, până la valori de 10 12 /cm 3 , de exemplu, în ape fecalo-menajere.
Bacteriile se pot întâlni la adâncimi mari în apa mărilor şi oceanelor, în ape termale. Existenţa în aer a bacteriilor este temporară şi prin intermediul curenţilor de aer sunt răspândite la distanţe foarte mari. Din aer, sunt antrenate din nou în sol prin intermediul precipitaţiilor atmosferice.
Bacteriile pot avea forme extrem de diferite. Astfel pot avea formă sferică (coci), de virgulă (vibrioni), de bastonaşi (bacili), sau de spirală (spirili).
Rolul bacteriilor în natură şi industrie. Bacteriile au un rol imens în transformarea compuşilor macromoleculari în compuşi simpli, prin mineralizarea materiei organice nevii, contribuind astfel la realizarea naturală a circuitului unor elemente de importanţă vitală: carbon, azot, sulf, fosfor, fier ş.a. Fără activitatea bacteriilor agenţi ai putrefacţiei - “pământul s-ar transforma treptat într-un uriaş cimitir”.
În industria alimentară bacteriile lactice sunt folosite la fabricarea produselor lactate, a brânzeturilor, în industria panificaţiei, la conservarea legumelor, măslinelor, furajelor verzi etc., bacteriile propionice sunt utilizate la fabricarea brânzeturilor tip schwaitzer, iar bacteriile acetice la fermentaţia alcoolului etilic - obţinerea industrială a oţetului.
Pe căi biotehnologice s-au obţinut cu ajutorul bacteriilor:
- enzime, proteine, aminoacizi, acid lactic, acid acetic, solvenţi (acetonă, alcool izopropilic, alcool butilic);
- hormoni - insulina produsă de un mutant de Escherichia coli;
- îngrăşăminte biologice - Azotobacter;
- insecticide biologice - Bacillus thuringiensis;
- antibiotice - Streptomyces sp.
- vitamine - de ex. vitamina B12 - Propionibacterium shermani
- Ca aspecte negative putem aminti:
- în industria alimentară - bacterii agenţi de alterare a produselor alimentare (acrirea berii, vinului, putrefacţia cărnii ş.a.);
- bacterii patogene - ingerare alimente contaminate - toxiinfecţii alimentare;
- bacterii patogene - pot să paraziteze organismele vii dând îmbolnăviri grave (tuberculoza, febra tifoidă, dizenteria, sifilis, bruceloza, antrax, ş.a.), bacterioze la plante.
Structura celulei bacteriene. Peretele celular. Fiecare bacterie este acoperită de un rigid perete celular, care este alcătuit din peptidoglucan. Peretele celular bacterian este considerat de mulţi un adevărat biopolimer. Acest perete îi dă celulei o formă şi înconjoară membrana citoplasmică, protejând-o de mediul înconjurător. Peretele celular bacterian de asemenea menţine integritatea celulară.
Acesta este atât de puternic, încât împiedică celula bacteriană să explodeze, atunci când presiunea osmotică dintre citoplasmă şi mediu este prea mare. Compoziţia peretelui celular diferă de la o celulă la alta, astfel alcătuind un factor important în analiza şi diferenţierea celulelor bacteriene. Astfel, în 1884, Hans Christian Gram, un fizician danez, a realizat o metodă de a diferenţia structurile peretelui celular.
Peretele celular gram pozitiv se colorează in violet prin tehnica Gram. Este gros, dar simplu structurat, unitatea de bază fiind peptidoglicanul, format din unităţi repetitive. Mureina este structura rezultată din unităţile de peptidoglicani cu ajutorul unei enzime: transpeptida. Are rolul de a menţine forma bacteriei (la presiuni de 2-2,5 atmosfere) in cazul in care bacteria işi pierde peretele sub acţiunea penicilinei. Sferoplastul (cu urme de perete celular) si protoplastul (fara perete celular) sunt fragili, usor de atacat de fagocite sau sub acţiunea variaţiei presiunii osmotice.
Membrana citoplasmică. Membrana citoplasmică procariotă constă dintr-un strat dublu de fosfolipide şi proteine, şi se aseamănă foarte mult cu membrana eucariotă. Aceasta este o barieră, ce le permite să interacţioneze selectiv cu mediul înconjurător. Membranele sunt asimetrice, constând fiecare din 2 părţi, fiecare parte constând din suprafeţe şi funcţii diferite.
Citoplasma. Nucleoidul, plasmidele şi cromozomii. Nucleoidul reprezintă regiunea citoplasmei, unde este concentrat ADN-ul cromozomal. Nu este un nucleu propriu-zis, ca la eucariote. Plasmide. Plasmidele pot păstra informaţia genetică adiţională. Ele se află în special în nucleoid. Informaţia păstrată de acestea nu este vitală, însă poate oferi avantaje selective în anumite circumstanţe. De ex: unele plasmide pot codifica anumite toxine, iar alte plasmide pot păstra informaţie genetică împotriva antibioticelor.
Cromozomi. Cromozomii bacterieni sunt în formă circulară şi păstrează informaţia genetică a bacteriilor.
Ribozomii şi alte structuri multiproteinice. Ribozomii sunt structuri prezente în orice celulă. Ribozomii bacterieni sunt similari celulelor eucariote, însă sunt mult mai mici şi au o altă structură. Unele antibiotice ar inhiba funcţionarea ribozomilor celulelor procariote, astfel cauzând moartea bacteriei.
Caractere morfologice coloniale
Mediul de bază pentru cultivarea bacteriilor - bulionul de carne lichid sau solidificat cu agar-agar (BCA). Prin reproducere pe mediu nutritiv solidificat ia naştere o colonie alcătuită din biomasa de celule rezultate prin sciziune din celula unică:
- colonii de tip S (smooth - neted lucios)
- colonii de tip R (rough - rugos, aspru, zbârcit)
- colonii de tip M - la bacterii producătoare de capsule, cu consistenţă gelatinoasă, mucoidă
Coloniile bacteriene pot prezenta în secţiune un profil lenticular, crateriform, triunghiular şi perimetru circular, dantelat sau cu ramificaţii rizoidale.
Pe BCA coloniile devin vizibile după 24-48 ore şi pot avea culori de alb, alb-crem, galben-auriu, oranj-roşu, albastru, fluorescenţă, caractere microscopice importante în identificare. Pe medii nutritive lichide bacteriile pot da tulburare şi sediment - bacteriile anaerobe sau formează la suprafaţa lichidelor voal caracteristic, fragil, cutat, gelatinos - bacteriile aerobe.
Bacteriile prezintă forme celulare foarte diversificate, dintre care forme de bază, monocelulare, precum şi forme derivate ale acestora ce rezultă în urma asocierii celulelor rezultate prin reproducere.
Dintre formele de bază fac parte următoarele:
- forma sferică - denumită coccus, în care sfera este perfectă, de exemplu la micrococi sau ovalară, de exemplu la enterococi, lanceolată la pneumococi şi reniformă la gonococi;
- forma bacilară - cilindrică , este cel mai frecvent întâlnită, este denumită şi bacterium. Formele pot fi drepte cu capete rotunjite - genul Enterobacter , cu capete retezate - genul Bacillus , fusiforme, măciucate - genul Corynebacterium , cu diametruvariabil - genul Mycobacterium ;
- dintre formele spiralate-elicoidale, specifice bacteriilor patogene, fac parte: forma vibrio , forma spirillum sub forma unor filamente rigide cu spire largi, forma spirocheta sub forma unor filamente flexibile cu mai multe spire;
- formele filamentoase sunt caracteristice bacteriilor miceliene cu habitatul în sol şi ape (actinomicete, chlamydobacterii etc.).
Caractere fiziologice generale ale bacteriilor
Bacteriile se caracterizează prin complexitate metabolică, cu o mare capacitate de adaptare, răspândire pe cele mai diverse medii, ca urmare a producerii de enzime care le permite utilizarea în nutriţie a compuşilor organici macromoleculari (protide, poliglucide, lipide).
În raport cu temperatura, bacteriile se dezvoltă într-un domeniu larg, între 10°C şi 90°C; majoritatea bacteriilor agenţi de alterare a alimentelor sunt bacterii mezofile şi dau alterări la temperatura camerei (bacterii de putrefacţie). Bacteriile în formă vegetativă sunt inactivate pe cale termică la temperaturi de pasteurizare, iar sub formă de endospori, la temperaturi de sterilizare.
În raport cu oxigenul majoritatea bacteriilor sunt aerobe (ex. bacterii acetice), care cresc în semiaerobioză (ex. bacterii lactice), dar un grup restrâns de bacterii sunt adaptate să crească în strictă anaerobioză (ex. genul Clostridium ).
Bacteriile se pot dezvolta în domeniul de pH1-11 cu zone optime la valori acide pentru bacterii acidotolerante (bacterii acetice, lactice) sau la valori neutre pentru bacterii de putrefacţie.
Clasificarea generală a bacteriilor
Pentru identificarea unei specii sunt uneori necesare 40 până la 100 de teste.
Criteriul de bază - afinitatea tinctorială; familia ca unitate taxonomică conţine acelaşi tip de bacterii: Gram pozitive sau Gram negative. Criterii fiziologice: teste de asimilare sau fermentare a glucidelor, relaţia faţă de oxigen, temperatură, pH, rezistenţă la inhibitori ş.a.
Clasificarea de bază - Bergey (1952) - bacteriile sunt grupate în 10 ordine şi 47 familii - clasificare reluată în 1984 după criterii morfologice - în 33 secţiuni.
Regnul procariotae - două diviziuni:
- Scotobacteria - bacterii care folosesc pentru creştere şI multiplicare energia rezultată din reacţii chimice: Clasa bacteria, Clasa actinomyces, Clasa molicutes
- Photobacteria -bacterii ce conţin pigmenţI celulari similari clorofilei şI care pot folosi energia luminoasă în procese de biosinteză celulară
Clasa BACTERIA
Ordinul pseudomonadales - bacterii Gram negative cu habitat în sol şi ape, aerobe, nesporulate.
Familia Pseudomonadaceae - genuri:
- Pseudomonas - bastonaşe tipice, răspândite pe produse vegetale, carne, pui; dau alterarea produselor refrigerate
- Acetobacter - bacterii acetice, produc oxidarea alcoolului etilic - la fabricarea acidului acetic de fermentaţie
- Xanthomonas - dau alterări ale legumelor, produc un polimer - xanthanul
- Zymomonas - bacterii care pot produce fermentarea glucidelor cu formarea de alcool; sunt folosite la obţinerea alcoolului carburant din materii celulozice.
Familia Nitrobacteriaceae , cu genurile Nitrobacter şi Nitrosomonas care produc oxidarea compuşilor cu azot rezultaţi din putrefacţie, cu transformarea azotului amoniacal în azotiţi şi azotaţi, formă asimilabilă de către plante.
Familia Thiobacteriaceae - produc oxidarea compuşilor cu sulf. Bacterii din genul Thiobacillus - agenţi ai coroziunii biologice.
Familia Spirillaceae - produc degradarea celulozei în condiţii aerobe. Bacterii ale genului Cellulomonas - prelucrarea deşeurilor de hârtie pentru obţinerea de proteină bacteriană folosită în scop furajer.
Ordinul EUBACTERIALES - bacterii propriu-zise, foarte răspândite ce cuprind bacterii în formă de coccus, bacterium şi forme derivate prin sciziune
Familia Achromobacteriaceae - bacterii nesporulate Gram negative, produc putrefacţia:
- g. Achromobacter - produc prin degradarea proteinelor amine biogene toxice, sunt bacterii aerobe, produc alterarea produselor refrigerate
- g. Alcaligenes - sunt întâlnite în lapte, carne, pui, peşte şi materii fecale
- g. Flavobacterium - se prezintă sub formă de bastonaşe, produc un pigment galben-roşu şi alterarea produselor la refrigerare.
Familia Azotobacteriaceae - bacterii care folosesc azotul atmosferic în nutriţie, au rol în circuitul natural al azotului şi pentru obţinerea de îngrăşăminte biologice - Azotobacter chroococcum
Familia Bacillaceae - bacterii sub formă de bastonaşe, Gram pozitive, producătoare de endospori:
- g. Bacillus - 25 specii - bacterii de putrefacţie aerobe,anaerobe; unele specii selecţionate se folosesc pentru obţinerea de enzime: amilaze, proteaze, glucanaze
- g. Clostridium - 93 specii - bacterii de putrefacţie anaerobe, producătoare de toxine, bacterii butirice, bacterii producătoare de solvenţi
- familia Enterobacteriaceae - bacterii Gram negative, nesporulate, aerobe/facultativ anaerobe, patogene/facultativ, cu habitatul în tractul digestiv
- g. Escherichia - bacterii de putrefacţie, facultativ patogene (agenţi ai gastroenteritelor) se pot înmulţi în produse alimentare, pot produce toxine E.coli - indicator sanitar pentru verificarea condiţiilor de igienă la fabricarea produselor alimentare
- g. Enterobacter - face parte din microflora intestinală
- g. Proteus - bacterii de putrefacţie, mobile, aerobe, produc alterarea cărnii, a ouălor păstrate la temperatura camerei
- g. Shigella - bacterii enteropatogene ( agentul dizenteriei)
- familia Lactobacillaceae - bacterii lactice Gram pozitive, nesporulate, facultativ anaerobe, sub formă de bacili (bastonaşe subţiri sau sub forme derivate de la coccus.
- g. Streptococcus - cuprinde bacterii sub formă de streptococi. O parte din speciile genului au trecut în g. Lactococcus , folosite drept culturi starter în industria laptelui
- g. Lactobacillus - bacterii lactice acidotolerante, folosite în industria laptelui şi pentru conservarea prin murare a produselor vegetale
- g. Pediococcus - bacterii lactice sub formă de tetrade. Pot produce acrirea berii
- g. Leuconostoc - bacterii lactice heterofermentative, agenţi de alterare a sucurilor, siropurilor de zahăr ş.a. Pot produce biosinteza dextranului
Familia Micrococcaceae - bacterii Gram pozitive cu forma coccus sau forme derivate prin sciziune
- g. Micrococcus - bacterii aerobe/anaerobe,de putrefacţie
- g. Sarcina - bacterii aerobe de putrefacţie
- g. Staphylococcus - bacterii facultativ patogene; produc enterotoxine şi sunt agenţi ai intoxicaţiilor alimentare
Familia Propionibacteriaceae - bacterii nesporulate Gram pozitive, produc fermentaţia propionică.
- g. Propionibacterium - bacterii folosite la fabricarea brânzeturilor cu pastă tare şi desen pentru obţinerea vitaminei B12.
CLASA ACTINOMYCES
Ordinul ACTINOMYCETALES - bacterii filamentoase, Gram pozitive, saprofite sau facultativ patogene, folosite industrial pentru obţinerea de substanţe biologic active
Familia Mycobacteriaceae - bacterii patogene - Mycobacterium tuberculosis (agentul tuberculozei) şi Mycobacterium leprae
Familia Actinomycetaceae - bacterii patogene pentru animale şi plante. Au rol în formarea humusului şi la închiderea la culoarea solului.
Familia Streptomycetaceae - bacterii filamentoase:
- g. Streptomyces - numeroase specii sunt folosite pentru obţinerea de antibiotice (tetracicline,streptomicina, cloramfenicol etc.) sau pentru enzime (glucozizomeraze).
Ordinul RICKETSIALES - cuprinde bacterii obligat parazite ale insectelor, transmisibile prin înţepături la animale şi om.
- g. Ricketsia - cu sp. R. prowazecki - agent al tifosului eczantematic
- g. Coxiella - bacterii patogene, produc febra Q hemoragică. C. bruneti se poate transmite prin lapte