Replicarea virusurilor
- Detalii
- Categorie: Microbiologie
- Accesări: 7,503
Procesul de replicare virală se realizează întodeauna numai în interiorul celulei gazdă pe care, virusul odată pătruns, o obligă prin informaţia genetică transmisă să sintetizeze componentele virale în dauna sintezei propriilor constituienţi. Se poate afirma că virusurile sunt multiplicate de celula-gazdă. Replicarea particulelor virale se desfăşoară sub forma unui ciclu, care începe cu fixarea acestora pe celula gazdă şi se termină cu eliberarea particulelor virale.
Principalele faze ale acestui ciclu sunt:
- fixarea şi adsorbţia pe celula gazdă.
- pătrunderea
- eclipsa
- sinteza componentelor virale
- maturarea
- eliberarea virusului.
Fixarea şi adsorbţia pe celula gazdă
Această etapă este posibilă întru-cât la suprafaţa membranei celulare există, în mod normal, constituienţi celulari denumiţi receptori de virus. Aceştia, din punct de vedere chimic, au compatibilitate cu structurile chimice din capsida virală sau pericapsidă. Desigur, nu orice tip de celulă posedă receptori pentru orice tip de virus, astfel că fixarea şi adsorbţia virusurilor pe celule nu este întâmplătoare şi depinde de afinitatea dintre celulă şi virus, afinitate denumită tropism. În absenţa receptorilor specifici celula gazdă este denumită celulă nepermisivă.
Există mai multe tipuri de tropism şi anume:
- tropism de regn ( vegetal, animal) –virusuri fitotrope (au acces numai la celule vegetale), virusuri zootrope (acces la celule animale), virusuri bacteriotrope (au acces la bacterii, bacteriofagii),
- tropism legat de specie, existând virusuri care parazitează o singură specie (ex:virusul peste porcine, virusul anemiei infecţioase ecvine) şi virusuri cu tropism pentru mai multe specii (ex:virusul turbării).
- tropism legat de tipul de celulă, existând virusuri neurotrope, virusuri epiteliotrope, virusuri pneumotrope, virusuri pantrope (tropism pentru celule diferite ).Privită în dinamică această etapă se desfăşoară în două faze:
- prima,numită faza reversibilă, se realizează datorită existenţei unei atracţii ionice dintre virus şi celulă.
- a doua, numită faza ireversibilă, se realizează doar atunci când există tropism între virus şi celulă.
Pătrunderea (penetraţia) virusului în celulă
În această fază se realizează infectarea celulei cu virus datorită pătrunderii acestuia prin peretele celular.Procesul poartă denumirea de viropexie. Acest proces activ se poate desfăşura prin două modalităţi:
1.Endocitoză, specifică virusurilor nude,iar mecanismul este asemănător procesului de fagocitoză.La nivelul membranei celulare se formează o invaginaţie care include virusul iar ulterior membrana celulei se reface.Învelişul viral va fi lezat de enzimele citoplasmatice eliberându-se genomul viral.
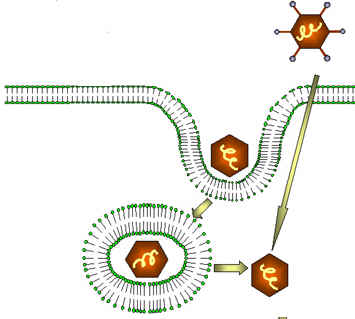
Figura 1 Pătrunderea în celulă prin endocitoza
2.Fuziune, specifică virusurilor acoperite, reprezintă pătrunderea directă a virusului prin peretele celular, la locul de fixare. Spiculii existenţi au rolul unor enzime, glicoproteaze care vor liza membrana celulară formând un canal prin care virusul trece în materia citoplasmatică.

Figura 2 Pătrunderea în celulă prin fuziune
Eclipsa sau decapsidarea
După pătrunderea particulei virale în celula gazdă într-un interval de una – câteva ore, are loc modificarea structurii morfologice a virisului prin distrugerea capsomerelor, sub acţiunea proteazelor celulare. Are loc eliberarea acidului nucleic, astfel că genomul viral devine liber denumit şi virus vegetativ. Acesta este inobservabil, fiind aparent dispărut ( fără identitate), de unde şi denumirea de eclipsă.
Unele virusuri pot rămâne în această stare mascată un timp îndelungat, sub formă de provirus, urmată apoi de faza de multiplicare activă. În schimb la alţii faza de eclipsă durează puţin (1-2 ore ) urmată apoi de faza de multiplicare activă. La unele virusuri genomul viral liber rămâne în citoplasmă iar la altele acesta pătrunde în nucleu.
Biosinteza componentelor virale
Acidul nucleic viral (ADN sau ARN) conţine informaţia genetică necesară replicării lui, inclusiv sintezei proteinelor virale. În general replicarea particulelor virale reuneşte trei aspecte comune tuturor grupelor de virusuri şi anume:
- replicarea genomului viral şi producerea de genomi progeni.
- transcripţia sau copierea informaţiei prin sinteza de ARN-m .
- translaţia sau decodificarea mesajului la nivelul ribozomilor celulari pentru sinteza proteinelor virale.
Astfel că celula gazdă pune la dispoziţia virusului, pentru replicare şi biosinteza proteinelor virale, materialul plastic şi energetic necesar: aminoacizi, nucleotide, enzime, ARN-t (transfer), ribozomi, iar virusul participă doar cu informaţia genetică cuprinsă în genomul viral, care indică ce trebuie realizat.
Maturarea particulelor virale
Constituienţii virali odată formaţi se vor asambla într-un tot unitar formând noi particule virale şi anume în jurul moleculelor de acid nucleic ,se dispun capsomerele într-o ordine şi simetrie foarte precisă.
Asamblarea poate avea loc în citoplasmă, la marea majoritate a ribovirusurilor, sau în nucleul celulei.
Eliberarea virusurilor
Particulele virale nou formate se eliberează din celula gazdă, existând mai multe modalităţi:
Liza celulei gazdă, adică prin distrugerea acesteia şi eliberarea întregi cantităţi de material viral. Acest mod de eliberare apare la virusurile foarte mici, nude şi sunt considerate virusuri agresive.
Înmugurire când la nivelul membranei celulare se formează muguri (excrescente) în care pătrund virionii şi care ,,împrumută” material de la nivelul membranei celulare pentru sinteza pericapsidei. În continuare incluşi în aceste excrescenţe virionii se detaşează de celula gazdă. Astfel, prin acest mod de eliberare celula gazdă nu este distrusă în totalitate, iar membrana celulară se poate reface.
Celula eliberează virioni treptat şi continuu,iar această posibilitate de punere în libertate a materialului viral este specifică virusurilor de dimensiuni mari şi acoperite care sunt considerate atenuate.
Durata ciclului de multiplicare este diferită, fiind mai redusă la ribovirusuri într-cât ARN-ul viral are rol şi de ARNm, şi cel mai adesea este monocatenar.
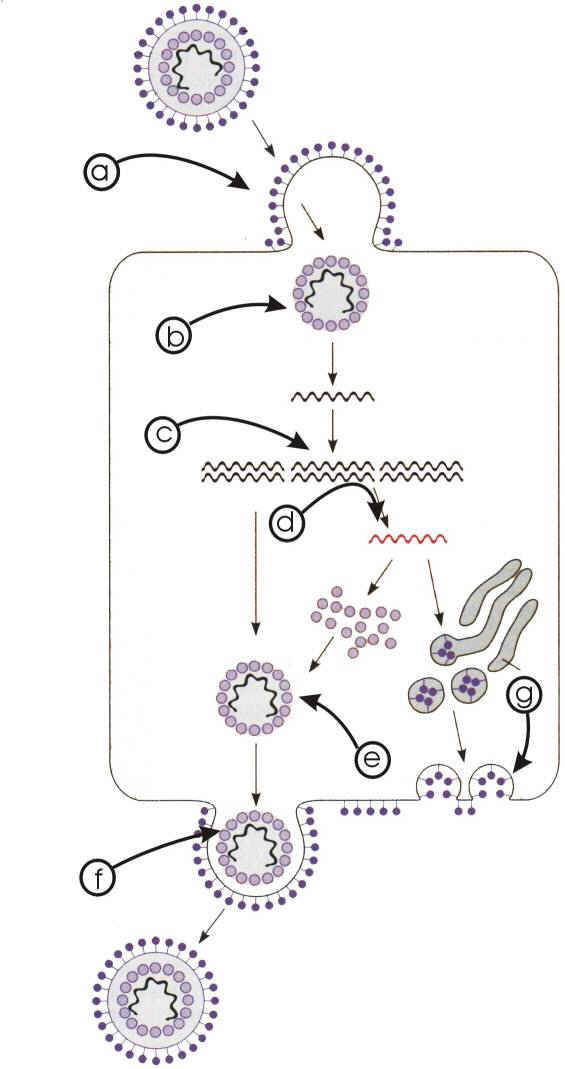
Figura 3 Schema multiplicării virale
- a– pătrunderea
- b– decapsidarea
- c–replicarea genomului
- d–transcripţia şi translaţia
- e– maturarea
- f– eliberarea
- g–biosinteza pericapsidei
Relaţia virus - celulă gazdă parazitată
Între virus şi celula gazdă infectată,după modul cum se realizează replicarea virală, există trei categorii de interelaţii şi anume.
Ciclul litic (citocid, citolitic) este caracteristic virusurilor agresive la care apar relaţii de antagonism acut cu efecte distructive asupra celulei gazdă datorită replicării exagerate urmată de distrugerea în totalitate a celulei gazdă parazitate. Particulele virale nou replicate şi eliberate atacă noi celule iar cele iniţial infectate nu mai produc material viral.
Virusurile care au la bază un asemenea ciclu sunt în general capabile să declanşeze boli grave,cu evoluţie acută şi transmisibile pe cale orizontală. În cursul acestor boli se pun în libertate,în organism şi în mediul înconjurător,cantităţi mari de virus printr-o serie de secreţii şi excreţii bogate în particule virale.(ex: febra aftoasă, gastroenterita infecţioasă a căţeilor,exantemul veziculos, virusurile fagice,etc.). Durata unui ciclu litic este variabilă,între 3o minute (pentru unii fagi) şi 5-20 ore (pentru virusurile patogene zootrope).
Ciclul necitolitic (persistent, endosimbiotic) este caracteristic virusurilor atenuate care au o rată mai redusă de multiplicare şi ca atare sunt mai puţin virulente. Celula infectată poate să supravieţuiască timp îndelungat, cu producerea de virioni maturi, compleţi (infecţie productivă) sau de componente virale (acid nucleic,capsidă-deci genom viral parţial exprimat-infecţie abortivă). În cadrul infecţiilor abortive virusurile incomplete pot fi maturate prin suprainfecţii cu ale virusuri numite virusuri helper.
Virusurile care au la bază un asemenea ciclu au o relaţie persistentă în timp cu celula gazdă iar infecţiile produse sunt lente,cronice, cu evoluţie progresivă şi modificări degenerative la nivelul celulelor gazdă.Transmiterea acestor infecţii virale se realizează tot pe cale orizontală (ex: boala Aujeszky, boala lui Marek,etc).
Ciclul lizogenic (integrat neproductiv) este caracteristic virusurilor temperate la care acidul nucleic viral pătrunde în genomul funcţional al celulei gazdă şi transcrie copii ADN, sub formă de provirus. Rămâne în această stare de latenţă (provirus) şi coexistă cu funcţiile normale ale ciclului, fără perturbări.În urma multiplicării celulare fiecare descendent va primi în zestrea sa genetică şi genomul viral (transmitere pe cale verticală ; ex: leucemiile la om şi leucozele la animale, anemia infecţioasă ecvină, SIDA, etc):
Virusurile latente pot rămâne în stare mascată fără a se multiplica timp îndelungat, dar când intervine un factor declanşator al multiplicării virale, se poate ajunge la infecţii acute virale extrem de grave.
De asemenea virusurile latente pot determina transformări ale celulei gazdă, sub raport morfo-funcţional şi mult mai grav, genetic. Nu mai există un control al creşterii şi multiplicării celulelor, survenind astfel modificări de tip oncogen.
Incluziile virale
Sunt formaţiuni omogene sau granulare, de forme şi dimensiuni variate, care apar în citoplasma sau nucleul celular infectate cu anumite virusuri şi sunt vizibile şi în câmpul microscopului optic.
Aceste incluzii reprezintă locul unde se sintetizeză componentele virale sau locul unde se asamblează particulele virale.Pentru unele virusuri aspectul incluziilor virale este caracteristic sau chiar patocnomomic(specific) şi au denumiri caracteristice pentru fiecare virus, de regulă poartă numele celui care le-a descoperit (ex:incluzii Babeş-Negri,în turbare, incluzii Bollinger în variolă,incluzii Rubarth în hepatita infecţioasă a câinelui).
Interferenţa virală
Interferenţa virală reprezintă procesul de inhibare a replicării unui virus de către un alt virus care a venit în contact înainte cu celula respectivă. Ea este rezultatul blocării ciclului normal de replicare a unui virus de către un alt virus, prin includerea unei rezistenţe celulare sau prin inducerea sintezei de interferon, capabil să înhibe producţia virusului de suprainfecţie. Virusul care declanşează interferenţa se numeşte virus inductor sau interferent, iar celălalt,virus interferat sau exclus.
Interferonul, descoperit în 1957, reprezintă o clasă de proteine sintetizată de celulele animalelor vertebrate infectate cu un virus şi care transmiţându-se altor celule mediază un mecanism de apărare faţă de agresiunea virală. Interferonul este eliberat de celula infectată şi difuzează în spaţiile extracelulare. Acţiunea protectoare a interferonului se exercită şi asupra altor tipuri de celule, de la alte specii decât a celei producătoare şi fără o sensibilitate virală.
Este cunoscut faptul că toate tipurile de celule pot produce interferon, însă unele produc cantităţi însemnate,ca de exemplu:celulele S.R.E., limfocitele, celulele splenice, celule hepatice, cele pulmonare, celule timice, etc.
Mecanismul de acţiune al interferonului nu este direct, virulicid, ci prin intermediul unei proteine denumită ,,proteină inhibitoare a translaţiei”.Prin intermediul acestei proteine celula este capabilă să facă distincţie între ARNm viral şi cel propriu, blocând astfel replicarea virală. Interferonul acţionează şi asupra virusurilor oncogene latente, împiedicând şi inhibând creşterea tumorală.
Totodată interferonul difuzând la celulele sănătoase învecinate înaintea virusului, le transmite acestora rezistenţa antivirală, împidicând infecţia.Astfel interferonul este utilizat în medicină în tratamentul unor boli virale şi în unele boli canceroase.