Introducere în fiziologia sistemului nervos
- Detalii
- Categorie: Anatomie si fiziologie
- Accesări: 10,329
Unitatea funcţională a sistemului nervos este reflexul realizat prin neuron. Un neuron este format din corp (soma) sau pericarion şi prelungiri denumite dendrite şi axoni (cilindracşi). Axonii sunt prelungirile prin care excitaţia pleacă, iar dendritele, acelea care vin la som. Forma corpului celular poate fi stelată, rotundă, piramidală, fuziformă, ovalară, piriformă etc.
După numărul prelungirilor pe care le prezintă şi după felul în care pornesc acestea din corpul celulei nervoase neuronii se clasifică în neuroni unipolari, pseudounipolari, bipolari şi multipolari. Neuronii unipolari nu prezintă decât axonul, polul receptiv fiind difuz, excitaţiile fiind recepţionate prin toată suprafaţa pericarionului. Astfel de neuroni sunt rari şi intră în structura retinei (celule amacrine).
Neuronii pseudounipolari sunt caracterizaţi prin aceea că din corpul lor pleacă o singură prelungire, dar după un traiect scurt se bifurcă în două ramuri ce reprezintă dendrita şi axonul; se găsesc în ganglionii spinali. Neuronii bipolari prezintă un axon şi o singură dendrită care pleacă de obicei din puncte opuse.
Astfel de neuroni se găsesc în retină, mucoasa olfactivă, în ganglionii lui Corti şi Scarpa. Neuronii multipolari prezintă un axon şi numeroase dendrite care pornesc de pe toată suprafaţa pericarionului.
Majoritatea neuronilor de acest tip se găsesc în toate segmentele sistemului nervos: celulele piramidale din cortexul cerebral, celulele radicaluare motorii din coarnele anterioare ale măduvei spinării, celulele din ganglionii vegetativi extranevraxiali.
Din punct de vedere funcţional neuronii pot fi motori, senzitivi şi de asociaţie. Neuronii motori sau efectori, de obicei sunt mari, multipolari, cu axonul lung ce se termină în organele efectoare (muşchi, glande). Aşa sunt celulele piramidale din scoarţa cerebrală, motoneuronii din coarnele anterioare medulare, celule Purkinje din scoarţa cerebeloasă. Neuronii motori pot somatomotori şi visceromotori.
Neuronii senzitivi sau receptori au proprietatea de a primi excitaţiile recepţionate de celulele senzoriale din mediul extern sau intern. Dendritele acestor neuroni senzitivi se distribuie la diferiţi receptori externi sau interni. Exemple de astfel de neuroni sunt neuronii pseudounipolari senzitivi, somatici şi vegetativi din ganglionul spinal, neuronii senzitivi din coarnele posterioare ale măduvei sau neuronii din ganglionii Corti şi Scarpa etc. Neuronii de asociaţie (intercalari), fac legătura între neuronul senzitiv şi cel motor. Sunt de dimensiuni mici, multipolari şi se găsesc în toate etajele sistemului nervos.
Proprietăţile fiziologice ale neuronului
Atât prelungiri care aduc impulsuri la soma neuronală, cât şi cele care le transmit la aceasta formează fibrele nervoase. Ele pot avea un diametru mai mare sau mai mic, pot fi mielinizate sau nemielinizate şi conduc impulsul cu o viteză în funcţie de aceste particularităţi. Fibrele nervoase intră în componenţa nervilor şi pe baza caracteristicilor menţionate au fost împărţite în diferite categorii (vezi tabelul. 1).
Tabel 1 - Clasificare fibrelor nervoase. În paranteză e trecută clasificarea numerică pentru fibrele senzoriale
| Tipul de fibre | Diametrul Fibrei μ | Grosimea | Viteza de conducere m/s | Originea şi funcţia | |
| A | α | 10-20 | groase | 60-120 | Proprioceptori, terminaţii anulo-spirale (I a), organul tendinos Golgi (Ib) |
| β | 7-15 | Subţiri | 40-90 | Fibre pentru contact şi presiune | |
| γ | 4-8 | Subţiri | 30-45 | Fibreγ - eferente ale fusului muscular | |
| δ | 2,5-5 | Subţiri | 15-25 | Receptori termici şi nociceptivi (III) | |
| B | 1-3 | Mai subţiri | 3-15 | Fibre preganglionare vegetative | |
| C | s | 0,3-1,2 | Foarte subţiri | 0,7-2,3 | Fibre eferente postganglionare simpatice |
| d | 0,3-1,3 | Foarte subţiri | 0,6-2,0 | Fibre eferente din rădăcina doprsală, pentru durere (IV) | |
Proprietăţile specifice neuronului sunt excitabilitatea şi conductibilitatea, proprietăţi generale ale tuturor ţesuturilor vii.
Excitabilitatea
Excitabilitatea este proprietatea celulelor de a răspunde la un stimul. Este o proprietate generală a materiei vii. Nervii, care sunt pachete de axoni şi dendrite, sunt dotaţi în consecinţă cu excitabilitate. Excitabilitatea a fost studiată cu ajutorul stimulilor electrici care pot fi uşor dozaţi ca intensitate şi timp. În acest scop se folosesc curenţii continui (galvanici) sau alternativi.
Excitabilitatea a fost caracterizată prin determinarea unor raporturi între stimul şi răspuns. Cei mai importanţi parametri ai excitabilităţii astfel studiate sunt: reobaza, timpul util, cronaxia, climaliza şi labilitatea.
Curent de bază, de prag sau liminar este intesitatea minimă de curent care produce un răspuns. Prin reobază se stabileşte pragul excitabilităţii sau intensitatea curentului liminar (fig. 1).

Figura 1 Curba excitabilităţii în funcţie de timp
Ea poate fi, ca şi alţi parametri, urmărită pe un preparat neuromuscular izolat, pe un nerv izolat sau pe organismul întreg, răspunsul fiind apreciat după contracţia musculară. În acest din urmă scop se stimulează pe suprafaţa cutanată puncte situate în apropierea unui câmp motor. O valoare mică a reobazei indică o excitabilitate crescută şi invers. Reobaza nervilor care inervează flexorii (muşchii albi) este mai mică decât a extensorilor (muşchii roşii), primii având deci o excitabilitate mai mare.
Timpul util este timpul minim necesar ca un curent de intensitatea reobazei să provoace un răspuns. El prezintă însă variaţii mari (2-20 ms) la modificări mici de curent şi, în consecinţă, are o valoare informativă redusă.
Cronaxia este timpul minim necesar pentru ca un curent dublu faţă de reobază să producă un răspuns din partea ţesutului, respectiv a nervului. Pentru caracterizarea excitabilităţii, determinarea cronaxiei este mai valoroasă decât a reobazei şi a timpului util, pentru că valoarea ei variază puţin la modificări mai mari de curent aşa cum rezultă din curba excitabilităţii în funcţie de timp, stabilită de Lapicque, care a şi introdus această noţiune. Valoarea cronaxiei este de 10-30 de ori mai mică decât a timpului util, variind între 0,1-0,6 ms.
Nervii muşchilor albi, ca şi aceştia din urmă, au o cronaxie mai mică şi, în consecinţă, o excitabilitate mai mare decât cei roşii. Acelaşi lucru rezultă şi din reobaza mai mică pe care o au primii. În trasmiterea sinaptică s-a atribuit o mare importanţă egalităţii dintre cronaxia nervilor şi a muşchilor corespunzători. Lapicque a presupus că transmiterea excitaţiei de la nerv la muşchi ar fi condiţionată de acest izocronism. Cronaxia lor are într-adevăr o valoare apropiată. Dacă diferenţa dintre ele are o valoare dublă (heterocronism), transmiterea impulsului nervos nu mai are loc. Se presupune că oboseala musculară s-ar putea datora şi acestui heterocronism.
Climaliza este alt parametru al excitabilităţii şi exprimă raportul dintre bruscheţea cu care se modifică intensitatea curentului şi excitaţie. Pentru a se obţine un răspuns la o anumită valoare a stimulului, de exemplu a reobazei intensitatea curentului prag trebuie atinsă cu o anumită viteză. Dacă ea creşte încet, nervul se acomodează şi răspunsul nu mai are loc. Excitarea liminară a nervului se obţine deci, cu o anumită bruscheţe minimă. La acelaşi curent, dacă intensitatea liminară este atinsă mai repede răspunsul apare mai prompt. Timpul de climaliză (timpul de creştere a curentului pentru obţinerea excitaţiei de prag) este de 2-3 ori mai scurt decât cronaxia.
Perioada refractară constă din inexcitabilitatea nervului după excitaţie, caracterizând şi alte ţesuturi. Ea a fost studiată însă, mai tardiv pe nerv. Perioada refractară se împarte în absolută şi relativă. În prima fază a perioadei refractere (0,5 ms), nervul nu mai răspunde la excitaţie, iar în a doua fază (2 ms), răspunde doar la un stimul mai puternic, nervul având deci un prag de excitabilitate mai mare. Faza de inexcitabilitate absolută a nervului corespunde cu durata potenţialului de vârf. Cea mai scurtă perioadă refractară o au nervii oculomotori. Faza de inexcitabilitate relativă este urmată de o fază de hiperexcitabilitatea.
Labilitatea este în funcţie de perioada refractară şi exprimă numărul maxim de stimul care pot fi însuşiţi de un ţesut, respectiv de nerv. Dacă frecvenţa stimulilor este mai mare decât labilitatea, o serie de stimuli sunt pierduţi. Optimul unei excitaţii constă din stimuli de frecvenţa labilităţii. Labilitatea diferiţilor axoni variază de la 100 la nivelul plăcii motorii până la 1000 la neuronul cortical. Cea mai mare labilitate (1500) o au celulele Renshaw din măduva spinării.
Parabioza reprezintă o modalitate particulară de răspuns a neuronilor atunci când asupra acestora acţionează stimuli care depăşesc capacitatea lor funcţională. Excitanţii puternici pot determina, mai întâi, o stare de excitaţie, urmând apoi o depolarizare intensă, modificând în felul acesta excitabilitatea nervului care poate deveni hipoexcitabil, chiar inexcitabil. Parabioza se desfăşoară în mai multe faze (hiperexcitabilitate, egalizare, paradaxolă, inhibiţie completă) şi este considerată ca un mecanism de apărare a neuronilor faţă de acţiunea unor excitanţi foarte puternici, ea fiind de fapt o inhibiţie de protecţie.
Procesul parabiotic este reversibil, după încetarea excitantului excitabilitatea nervoasă revine la normal, parcurgând în sens invers fazele prin care a trecut.
Conductibilitatea
Conductibilitatea nervilor constă în proprietatea lor de a transmite un impuls generat într-un anumit punct. Impulsul este transmis în ambele direcţii, aşa cum reiese din înregistrarea curenţilor de acţiune. Conducerea bilaterală este astfel una din proprietăţile fibrei nervoase; dar în organismul întreg impulsul nervos este transmis într-un singur sens datorită polarizării sale la nivelul sinapselor din sistemul nervos central.
Conducerea unilaterală în sensul unui arc reflex este denumită ortodromică, spre deosebire de conducerea în sens invers, adică antridromică. Transmiterea liniară a excitaţiei se datoreşte depolarizării zonei excitate. Transmiterea retrogradă este împiedicată de perioada refractară care se instalează după excitaţie. Dacă se plasează doi electrozi pe o fibră nervoasă excitată, ca şi pe alt ţesut muscular liniar, se înregistrează un curent de acăiune bifazic.
Viteza de transmitere a excitaţiei este în funcţie de dimensiunile fibrei şi de mielinizarea nervului. Este mai mare în fibrele mai groase şi în cele mielinizate (de 50 de ori). După viteza de transmitere a excitaţiei, ca şi după grosimea lor, nervii sunt împărţiţi în diferite categorii (tabela 1).
Viteza mare din fibra mielinizată se datoreşte transmisterii saltatorii a impulsului de la o strangulaţie Ranvier la alta, singurele zone excitabile în aceste condiţii. De la o strangulaţie Ranvier la alta, stimulul este transmis prin mediu, care este un bun conductor datorită compoziţiei electrolitice cu proprietăţi de conducere.
Stimulul sare intervalul dintre cele două ştrangulaţii, mielina fiind un izolator relativ (fig. 2). Se activează astfel, nodulul lui Ranvier. În axonii amielinici, excitaţia este transmisă din aproape în aproape, prin depolarizarea membranei prin curenţi locali Hermann (fig. 3), ceea ce explică viteza mai lentă de răspândire.
Conducerea influxului dealungul unui axon integru se face izolat (legea conducerii izolate). Prin lezarea axonului excitaţia este transmisă unui axon vecin.
Degenerescenţa walleriană, descrisă de Waller în secolul trecut, constă în degenerarea nervului izolat prin secţiune de somă, deci a segmentului distal (anterograd). Suturând capătul perific al nervului cu cel central, el se regenerează datorită funcţiei trofice a tecii fibroase care îl înconjoară. Această regenerare se poate produce şi dacă segmentul distal este suturat cu un alt nerv, ceea ce demonstrează plasticitatea sistemului nervos central.

Figura 2 Conducerea saltatoare

Figura 3 Curenţii Hermann de-a lungul unui axom mielinizat
În sistemul nervos central degenerescenţa axonului secţionat are loc în sens bilateral. De exemplu, secţionând traiectul piramidal la nivel medular, degenerescenţa se produce atât periferic (anterograd), cât şi central (retrograd). Urmărirea acestei degenerescenţe constituie o metodă histologică utilizată pentru precizarea diferitelor conexiuni.
Potenţialul de repaus şi cel de acţiune
Excitaţia şi propagarea stimulului dealungul nervilor sunt însoţite de fenomene electrice. Plasând doi electrozi pe suprafaţa unei fibre nervoase, în repaus, nu se remarcă nici o deosebire de potenţial. Dacă însă, unul pătrunde în citoplasmă, se constată că interiorul axonului are un potenţial negativ de 70 mv. Acesta este generat în repaus de distribuţia inegală a ionilor de Na şi K, datorită pompelor active care polarizează membrana. Prin stimulare, aceasta este depolarizată datorită unui influx de ioni de Na. Potenţialul de acţiune dispare rapid, membrana fiind repolarizată (fig. 4).

Figura 4 Potenţialul de acţiune
Potenţialul de repaus este restabilit iniţial prin efluxul ionilor de potasiu şi influxul celor de clor. Cheltuiala de energie necesară proceselor de polarizare şi depolarizare este furnizată de oxidarea tisulară din mitocondrii, prin metabolizarea unei importante cantităţi de glucoză (ţesutul nervos este cel mai sensibil la hipoglicemie).
Pentru a obţine o excitaţie care se propagă, intensitatea stimulului trebuie să aibă o valoare minimă (pragul excitabilităţii), care variază de la un tip de fibră la altul. Fibra nervoasă este excitată după legea tot sau nimic. Excitanţii subliminari produc modificări, nemanifestate însă, coborând sau ridicând pragul excitabilităţii, în funcţie de efectul lor de depolarizare sau de hiperpolarizare. Dacă depolarizarea atinge un anumit nivel critic (10-30 mV), ia naştere un potenţial de acţiune care se propagă sub formă de excitaţie.
Electrotonusul
Stimulii electrici subliminari modifică excitabilitatea în mod diferit la catod şi la anod. La catod se produce o acumulare de sarcini pozitive, intracelulare care reduc polarizarea de repaus. Această depolarizare creşte progresiv cu intensitatea stimulului, până când diferenţa de potenţial este redusă cu 15 mV, moment în care are loc procesul de activare cu apariţia potenţialului de acţiune.
La anod, o redistribuţie ionică inversă creşte valoarea potenţialului de repaus prin hiperpolarizare. Întreruperea curentului poate declanşa un curent de acţiune. Aceste modificări de polarizare produse de stimuli sub prag poartă denumirea de electrotonus, cele de la catod fiind denumite catelectrotonus, iar cele de la anod, anelectrotonus.
Transmiterea sinaptică
Impulsul nervos este transmis de la un neuron la altul cu ajutorul mediatorilor chimici, dintre care cel mai cunoscut este acetilcolina. Acţiunea ei durează puţin, datorită colinesterazei aflate la acest nivel şi care o inactivează rapid. Efectul stimulării poate fi prelungit împiedicând acţiunea acestei enzime cu ajutorul unor substanţe farmacodinamice numite anticolinesteraze (ex. ezerina). Alături de acetilcolină, ca mediator chimic sinaptic pot interveni şi alte subtanţe ca: acidul glutamic, noradrenalina, serotonina, histamina.
Transmiterea chimică la nivelul sinapsei explică întârzierea de 0,7 ms pe care o prezintă, ca şi marea fatigabilitate (oboseală), aşa cum rezultă din epuizarea transmiterii dacă excitaţia are o frecvenţă de 40-50 impulsuri/s. Sinapsele sunt foarte sensibile la hipoxie prin afectarea ATP-ului, generator de energie pentru sinteza mediatorului. Impulsul sinaptic este descărcat repetitiv, ceea ce se explică prin eliberarea treptată a mediatorului.
Mediatorul chimic este eliberat sub impulsul potenţialului de acţiune al axonului, care conţine la capătul sinaptic vezicule de acetilcolină. Acetilcolina sau ceilalţi mediatori chimici sunt eliberaţi în fanta sinaptică în cca. 0,6 ms de la stimulare, producând o depolarizare a somei neuronale postsinaptice (fig. 5). Când depolarizarea atinge nivelul critic, impulsul este transmis spre periferie prin axonul postsinaptic.

Figura 5 - Sinapsa neuro-neuronală
Potenţialul postsinaptic excitator
Dacă stimulul presinaptic este slab, în soma motoneuronului se produce o depolarizare, care apare ca un potenţial de acţiune redus, cu durata de cca. 20 ms. Acesta nu este urmat de o activare axonală, rămânând localizat. El este denumit potenţial postsinaptic excitator (PPSE) şi creşte proporţional cu intensitatea stimulului. PPSE este produs printr-un influx ionic în zona neuronală, provocat de depolarizarea membranei sinaptice; este monofazic şi proporţional cu intensitatea impulsurilor.
Când polarizarea somei neuronale este redusă la un nivel critic de 60 mV, apare o depolarizare bruscă. Acest nivel critic provoacă un potenţial de acţiune care se propagă dealungul axonului. Potenţialul intracelular ajunge astfel, de la -70 mV la +20 mV, fiind deci de 90 mV faţă de nivelul iniţial. Spre deosebire de PPSE, potenţialul de acţiune se supune legii tot sau nimic, nefiind neproporţional cu intensitatea excitaţiei.
Potenţialul postsinaptic inhibitor
Anumiţi stimului sinaptici în loc să producă o depolarizare a somei neuronale, produc hiperpolarizare până la -80 mV. Acest potenţial poartă denumirea de potenţial postsinaptic inhibitor (PPSI). El realizează o imagine în oglindă faţă de PPSE.
Soma neuronală este astfel inhibată, stimularea necesitând o excitaţie mai puternică. PPSI poate fi produs de exemplu prin stimularea unui nerv antagonist ipsilateral; este generat probabil de un transfer ionic prin porii somei neuronale, cu dimensiuni de cca. 5 Å suficiente pentru a permite trecerea pe K+ şi Cl-, dar nu de Na +. K+ Părăseşte astfel neuronul, concomitent cu influx de Cl-, care îi suplimentează sarcini negative.
Hiperpolarizarea este produsă de mediatori chimici inhibitori cu o compoziţie puţin cunoscută. Se atribuie un rol important în acest sens acidului gammaaminobutiric (GABA). Unii cercetătorii susţin că pe suprafaţa somei neuronale există zone inhibitoare astfel încât inhibiţia ar fi determinată de locul de aplicare a stimulului.
Inhibiţia neuronală produsă prin hiperpolarizare sau prin alte mecanisme este tot atât de importantă ca şi excitaţia.
Prin ea, răspunsul motor este modelat şi coordonat, realizând o corespondenţă între activitatea muşchilor antagonişti. S-au descris trei tipuri de inhibiţie neuronală: directă, presinaptică şi recurentă (fig. 6).
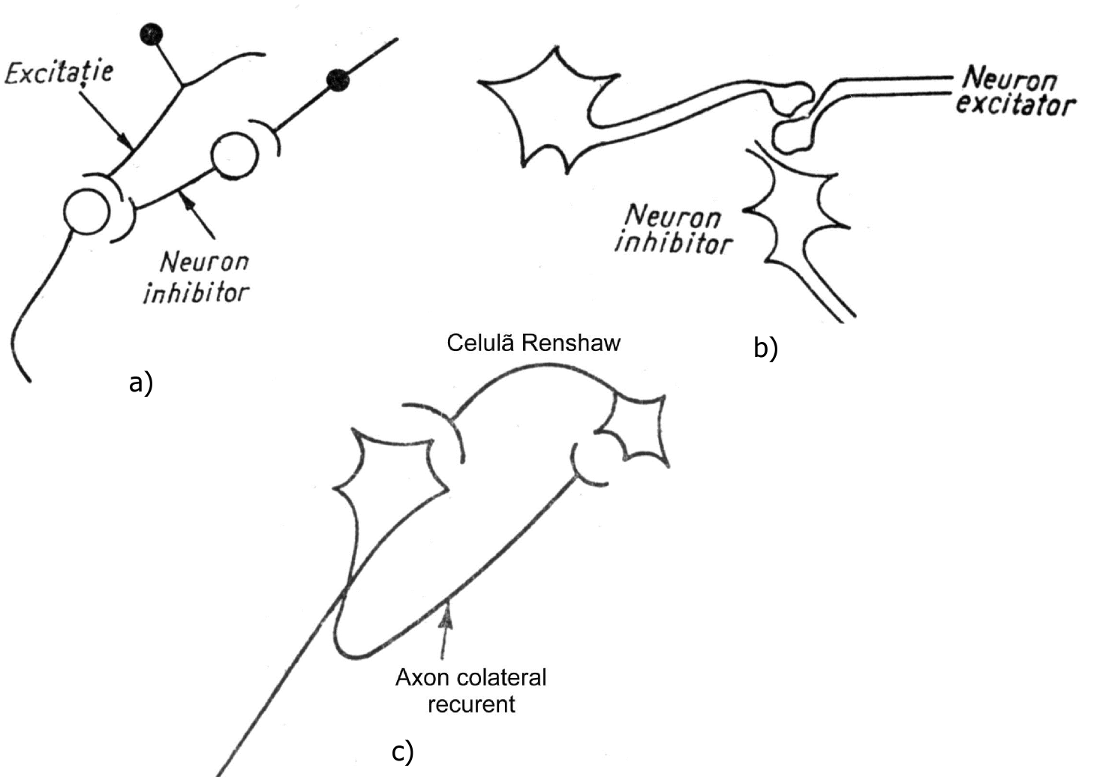
Figura 6 - a) Inhibiţie directă; b) Inhibiţie presinaptică; c) Inhibiţie de tip Renshaw
Inhibiţia directă constă din intercalarea unui neuron inhibitor care, prin mediator inhibitor, acţionează asupra unui neuron stimulat convergent.
Inhibiţia presinaptică diminuă descărcarea mediatorilor la nivelul sinapsei (cantitatea fiind proporţională cu intensitatea potenţialului de acţiune), aşa cum rezultă din scăderea PPSE. Ea este produsă de neuroni care acţionează pe terminaţiile celor excitatori sau pe axon.
Inhibiţia recurentă este mediată de celulele Renshaw, care sunt intercalate într-o ramificaţie axonală recurentă şi motoneuron. Stimulul se întoarce astfel la motoneuron, pe care îl inhibă, producând o hiperpolarizare.
Proprietăţile centrilor nervoşi
Numărul neuronilor care intră în alcătuirea organelor nervoase centrale este foarte mare. Impulsurile nervoase care apar la nivelul receptorilor sunt transmise neuronilor din centri nervoşi. La nivelul sinapselor centrale impulsurile vor trece la neuronii motori, fie direct, fie indirect prin intermediul neuronilor intercalari.
Centrii nervoşi, existenţi în diferite etaje nervoase, nu sunt izolaţi, ci între ei există numeroase legături. Datorită diferitelor procese de transmitere sinaptică, centrii nervoşi conferă impulsului un caracter diferenţiat şi nuanţat. Transmiterea excitaţiei prin centrii nervoşi se caracterizează printr-o serie de particularităţi.
Conducerea unilaterală a impulsului
După cum s-a văzut, în fibra nervoasă impulsul este transmis în ambele sensuri. În centrii nervoşi, transmiterea este însă unilaterală, fapt explicat prin mecanismul de eliberare a acetilcolinei la nivelul butonului terminal unde se depolarizează porţiunea de membrană subiacentă.
Întârzierea sinaptică
Perioada latentă a unui reflex este mai mare decât cea justificată de timpul necesar pentru stimularea receptorului, conducerea prin căile aferente şi eferente şi răspunsul organului efector. Perioada latentă mai lungă este explicată prin întârzierea centrală sinaptică, adică prin timpul necesar ca un stimul să fie transmis de la terminaţia axonală la corpul neuronal. La nivelul fiecărei sinapsei impulsul întârzie 0,5-0,7 ms. Cu cât centrul nervos este alcătuit dintr-un număr mai mare de neuroni, cu atât este mai mare întârzierea centrală sinaptică. În consecinţă, după durata perioadei latente se poate aprecia cu aproximaţie şi numărul neuronilor care formează centrul unui reflex.
Convergenţa şi divergenţa
La suprafaţa fiecărui neuron se termină un mare număr de prelungiri axonale. Fenomenul poartă denumirea de convergenţă. În acelaşi timp axonul unui singur neuron vine în contact prin terminaţiile sale cu un mare număr de corpuri neuronale printr-un fenomen denumit divergenţă (fig. 7).

Figura 7 - a) Convergenţa; b) Divergenţa
Se presupune că fiecare neuron primeşte aferenţe axonale de la cca. 100 de neuroni, sau mai mulţi şi trimite terminaţii axonale la cca. 100 de neuroni. Posibilităţile de interconexiune neuronală sunt deci practic infinite.
Postdescărcarea
Prin postdescărcare se înţelege faptul că, numărul impulsurilor din nervii eferenţi este mai mare decât al celor din fibrele aferente. În consecinţă, prin trecerea prin centrii nervoşi, stimulii sunt multiplicaţi. Acest proces poate fi explicat prin fenomenul divergenţei, în sensul că un stimul unic al nervului aferent este descărcat, prin numeroase ramificaţii axonale, într-un număr mai mare de neuroni motori eferenţi. Multiplicarea stimulului la nivelul centrului nervos este importantă pentru realizarea contracţiei tetanice, care este contracţia fiziologică a muşchiului striat.
Recrutarea
Prin recrutare se înţelege creşterea numărului de unităţi motoare stimulate în legătură cu creşterea intensităţii stimulului.
Inducţia simultană şi succesivă
Centrii nervoşi sunt sediul unor procese de coordonare care sincronizează activitatea muşchilor antagonişti. Coordonarea este realizată prin inducţie, care poate fi simultană sau succesivă. De exemplu, concomitent cu stimularea neuronilor care inervează extensorii unui membru se produce relaxarea flexorilor, care sunt antagonişti. Fenomenul se numeşte inducţie negativă simultană. Acesta este un proces de coordonare locală, probabil prin inhibiţie neuronală directă. Astfel, de exemplu, în decursul reflexului osteotendinos, impulsurile aferente sunt transmise neuronilor motori ai extensorilor, în timp ce este stimulat şi un neuron inhibitor intercalar care inhibă motoneuronii flexorilor (fig. 8).

Figura 8 - Conexiuni centrale probabile în coordonarea muşchilor antagonişti
După ce motoneuronii extensorilor au fost stimulaţi ei sunt inhibaţi probabil prin hiperpolarizare. Fenomenul se numeşte inducţie negativă succesivă. În acelaşi timp, flexorii, anterior inhibaţi, sunt stimulaţi, tot prin inducţie succesivă, dar pozitivă. Prin această interferenţă dintre excitaţia şi inhibiţia din centrii nervoşi sunt coordonate extensia şi flexia unui membru. Procese de inducţie succesivă survin şi la nivelul altor centri nervoşi care aferentează musculatura antagonistă, ca de exemplu în centrii nervoşi din trunchiul cerebral, care comandă mişcări de masticaţie, coordonând coborârea şi ridicarea mandibulei.
Iradierea
Iradierea este fenomenul prin care creşterea intensităţii stimulului activează o zonă progresiv, mai mare de neuroni centrali.
Organizarea funcţională a sistemului nervos
Sistemul nervos îndeplineşte rolul de reglare, coordonare şi integrare a funcţiilor tuturor organelor. El asigură în acelaşi timp unitatea organismului cu mediul înconjurător, realizând unitatea organism-mediu şi coordonează activitatea organelor interne.
Sistemul nervos central este în permanenţă informat prin stimulii recepţionaţi din afara şi din interiorul organismului, ceea ce face posibilă integrarea organismului în mediul înconjurător şi menţinerea constantă a mediului intern (homeostazia). Reglarea nervoasă a funcţiilor corpului se bazează pe activitatea centrilor nervoşi care prelucrează informaţiile intrate şi elaborează comenzi ce sunt transmise efectorilo. Fiecare centru nervos poate fi separat în două compartimente funcţionale:
- compartimentul senzitiv, unde sosesc informaţiile culese de la nivelul receptorilor;
- compartimentul motor, care transmite comenzile la efectori.
Deci fiecare organ nervos are două funcţii fundamentale: funcţia senzitivă şi funcţia motorie. La nivelul emisferelor cerebrale mai apare şi funcţia psihică. Separarea funcţiilor sistemului nervos este doar cu caracter didactic; în relitate nu există activitate senzitivă fără manifestări motorii şi viceversa, iar stările psihice derivă din integrarea primelor două.
Funcţiile fundamentale ale sistemului nervos sunt funcţia reflexă şi funcţia de conducere.
Funcţia reflexă
Reflexul reprezintă mecanismul fundamental de activitate a sistemului nervos. El reprezintă reacţia de răspuns a centrilor nervoşi la stimularea unei zone receptoare. Termenul a fost introdus în urmă cu 300 de ani de către matematicianul şi filosoful francez Rene Descartes. Răspunsul reflex poate fi excitator sau inhibitor.
La realizarea unui reflex participă cinci componente anatomice: receptorul, calea aferentă, centri, calea eferentă şi efectorul. Ele alcătuiesc împreună arcul reflex.
Receptorul este de obicei o celulă sau un grup de celule diferenţiate şi specializate în celule senzoriale (gustative, auditive, vizuale, vestibulare). Alţi receptori sunt corpusculii senzitivi alcătuiţi din celule, fibre conjunctive şi formaţiuni nervoase dendritice (receptorii tegumentari şi proprioceptorii). Uneori rolul de receptor îl îndeplinesc chiar terminaţiile butonate ale dendritelor (receptorul olfativ, receptorii dureroşi).
La nivelul receptorului are loc transformarea energiei excitantului în influx nervos. Fiecare receptor este specializat în transformarea unei anumite forme de energie din mediu (excitanţi sau stimuli) în informaţie nervoasă specifică (influxul nervos sau potenţialul de acţiune) cu amplitudine proporţională cu intensitatea excitantului. În acelaşi timp, fiecare receptor poate fi stimulat de orice formă de energie dacă depăşeşte cu mult intensitatea normală; astfel celulele vizuale pot fi excitate şi de energii mecanice mari (o lovitură cu pumnul în ochi provoacă senzaţii vizuale rudimentare).
Receptorii se pot clasifica după mai multe criterii.
Dupa localizare:
- exteroceptori, situaţi la nivelul pielii şi organelor de simţ, care culeg din mediul extern stimuli termici, dureroşi, tactili, olfactivi, vizuali, auditivi;
- proprioceptorii, situaţi la nivelul muşchilor, tendoanelor, articulaţiilor, periostului şi ligamentelor;
- interoceptori (visceroceptori) care culeg stimulii de la nivelul arborelui vascular şi al organelor interne (viscere).
După specializare:
- mecanoreceptori, sensibili la excitanţii mecanici;
- chemoreceptori, sensibili la acţiunea substanţelor chimice;
- osmoreceptori, sensibili la variaţiile presiunii osmotice;
- termoreceptori, sensibili la variaţiile de temperatură;
- voloreceptori, sensibili la variaţiile de volum;
- fotoreceptori, sensibili la energiile luminoase;
- algoreceptori, sensibili la stimulii nocivi, dureroşi.
Calea aferentă
Receptorii vin în contact sinaptic cu terminaţiile dendritice ale neuronilor senzitivi din ganglionii spinali sau de pe traiectul unor nervi cranieni. Variaţiile de potenţial de receptor produc depolarizări pasive în terminaţia dendritică care atingănd pragul critic descarcă potenţial de acţiune după legea "tot sau nimic" ce se propagă celulipet (aferent). Informarea corectă= a centrilor privind variaţiile energiei excitantului se face prin modulare de frecvenţă (modularea în amplitudine nu este posibilă din cauza legii tot sau nimic).
Ca urmare un potenţial de receptor de amplitudine redusă determină numai câteva potenţiale de acţiune pe secundă, în timp ce potenţiale de receptor mai ample induc zeci sau sute de potenţiale de acţiune pe secundă. Cea mai simplă cale aferentă este reprezentată de neuronul senzitiv spinal şi prelungirile sale.
Centrul
În cazul unui arc reflex elementar format din doi neuroni (unul senzitiv, celălalt motor) centrul reflex este reprezentat chiar de sinapsa dintre axonul neuronului senzitiv şi corpul neuronului motor (exemplu reflexul miotatic). Prin centrul unui reflex se înţelege totalitatea structurilor din sistemul nervos central care participă la actul reflex respectiv. Spre exemplu, centrii reflexelor respiratori se află în bulb, în punte precum şi în hipotalamus şi în scoarţa cerebrală.
Complexitatea şi întinderea unui centru este în funcţie de complexitatea actului reflex pe care îl efectuează. Centrul nervos care primeşte informaţia o prelucrează, o compară cu date din memorie, elaborează decizii şi controlează permanent modul de execuţie efectuând corectările necesare. La nivelul sinapselor din centrii reflecşi, transmiterea se face din nou prin modulare în amplitudine deoarece potenţialele postsinaptice nu mai respectă legea "tot sau nimic".
Calea eferentă este reprezentată de axonii neuronilor efectori (motori sau secretori). Cea mai simplă cale eferentă se găseşte la reflexele monosinaptice când este formată din axonul motoneuronului α. În cazul sitemului nervos vegetativ, calea efrentă este formată dintr-un lanţ de doi neuroni motori: un neuron preganglionar situat în coarnele laterale ale măduvei spinării sau într-un nucleu vegetativ din trunchiul cerebral şi un neuron postganglionar situat în ganglionii vegetativi periferici. De-a lungul căilor eferente informaţia circulă spre efectori din nou prin modulaţie de frecvenţă.
Efectorii
Principalii efectori sunt muşchiul striat, muşchiul neted şi glandele exocrine. În funcţie de fracvenţa potenţialelor de acţiune sosite pe axon, la nivelul plăcii motorii de exemplu, se vor sparge un anumit număr de vezicule cu acetilcolină, care va determina potenţiale postsinaptice de amplitudini diferite în funcţie de numărul de molecule de acetilcolină eliberate. Ca urmare, pe membrana fibrei striate apar zeci sau sute de potenţiale de acţiune pe secundă, producând contracţii de amplitudine şi forţe corespunzătoare comenzii centrale.
Controlul îndeplinirii comenzii
Arcul reflex cu cele cinci componente al sale reprezintă un model incomplet al desfăşurării activităţii reflexe. În ultimele decenii s-a descoperit existenţa unor noi circuite nervoase ce leagă centrii de organele receptoare. Prin intermediul acestora, centrii nervoşi pot regla pragul de excitabilitate al receptorilor şi implicit intensitatea stimulilor aferenţi. Un asemenea control se exercită asupra efectorilor musculari de către centrii motori extrapiramidali şi cerebrali.
În acelaşi timp de la nivelul efectorilor porneşte spre centrii un circuit recurent care îi informează asupra modului îndeplinirii comenzii (feed-back). Mecanismul de feed-back are caracter universal ce se întâlneşte în toate sistemele autoreglate. Comanda şi controlul exercitate de centrii nervoşi sunt de natură reflexă. În acest sens, centrii nervoşi nu sunt numai senzitivi sau numai motori, ci ei reprezintă centrii de integrare senzitivo-motorie. Răspunsul reflex poate surveni imediat după acţiunea stimulului sau poate întârzia minute, zile sau ani; în aceste cazuri informaţia ce intră în centrii este depozitată sub formă de memorie şi va fi actualizată în momentul elaborării răspunsului.
În raport cu efectorul asupra căruia se exercită preponderent comanda, sistemul nervos poate fi împărţit în:
- sistem nervos al vieţii de relaţie (somatic) care controlează efectorul muscular striat voluntar;
- sistem nervos al vieţii vegetative (de nutriţie) care controleazăn efectorul muscular neted şi glandular, coordonând activitatea organelor interne (viscere).
Este format din sistem nervos simpatic (care are ca mediator chimic noradrenalina şi adrenalina) şi sistem nervos parasimpatic (a cărei activitate este mediată chimic de acetilcolină).
Activitatea sistemului nervos somatic şi a celui vegetativ nu se desfăşoară separat ci într-o unitate perfectă; hipotalamusul, sistemul limbic şi scoarţa cerebrală integrează şi coordonează activitatea viscerelor, glandelor endocrine şi cea somatică. În felul acesta se realizează atât unitatea organismului în ansamblu cât şi unitatea organism-mediu.
Funcţia de conducere
Excitaţiile culese la nivelul suprafeţelor receptoare sunt conduse spre centrii, iar comenzile elaborate aici sunt conduse la periferie la organele efectoare. Întreg sistemul nervos apare ca un ansamblu imens de circuite funcţionale, bidirecţionale prin care informaţiile circulă necontenit.
Viteza de conducere a influxului nervos depinde de structura şi grosimea fibrelor nervoase. Fibrele cu diametru mare conduc mai repede decât cele înguste, iar pentru aceeaşi grosime fibrele mielinice conduc cu viteze mai mari decât cele amielinice. Viteza cea mai mare o au fibrele căilor proprioceptive (120 m/s) iar viteza cea mai redusă o au fibrele amielinice ca transmit stimulii dureroşi (0,5 m/s). Conducerea excitaţiei de la periferie spre centru se numeşte conducere afentă, iar a comenzilor de la centru la periferie se numeşte conducere eferentă.
Funcţia de conducere şi funcţia reflexă sunt strâns corelate, separarea lor se face numai în scopuri didactice.
Atât sistemul nervos somatic cât şi cel vegetativ au o componentă centrală (centrii nervoşi) situată în sistemul nervos central (măduva spinării şi encefalul) şi o componentă periferică (ganglioni şi trunchiuri nervoase), ce constituie sistemul nervos periferic.