Nucleul
- Detalii
- Categorie: Biologie celulară
- Accesări: 34,992

Nucleul celular este una dintre cele mai importante structuri ale celulei eucariote. Acesta funcționează ca un centru de comandă, controlând activitatea celulară și stocând materialul genetic. În acest articol, vom explora ce este nucleul celular, ce funcții are și de ce este esențial pentru viață.
Ce este nucleul celular?
Nucleul celular este un organit membranos prezent în celulele eucariote. Acesta conține ADN-ul (acidul deoxiribonucleic), materialul genetic care dictează structura și funcțiile celulei. În contrast, celulele procariote, cum ar fi bacteriile, nu au un nucleu organizat, ci un nucleoid în care este dispersat ADN-ul.
Istoria cercetării nucleului celular
Studiind la microscopul optic, Antony van Leeuwenhoeck [1702] pe sânge de broască, şi apoi Felice Fontana [1781] pe epidermă de anghilă, au fost primii care au menţionat în lucrările lor nucleul celular. În 1831, Robert Brown (pe Orchideae şi Asclepiadeae) şi Charles F. Brisseau de Mirbel (pe Marchantia) au descris nucleul în interfază, considerându-l un organit constant al celulei (ca o veziculă), format dintr-o membrană şi un conţinut lichid.
În 1882, Walther Flemming şi Edouard Strasburger au descris nucleul în timpul diviziunii celulare. În 1910 Thomas Hunt Morgan pus în evidenţă rolul genetic al nucleului, în 1953 James Watson şi Francis Crick au reuşit să schematizeze structura moleculei de ADN nuclear, iar în 1965 Jacques Monod, Francis Jacob şi Andre Lwoff au primit premiul Nobel pentru explicarea rolului genei la Escherichia coli sp.
În celulele procariote nu există un nucleu distinct, ci o singură moleculă circulară de ADN, cu aspect dispersat în interfază şi de cromozom unic în timpul diviziunii. Prin metode citochimice s-a evidenţiat o delimitare a genomului (material genetic) într-o structură numită nucleoid sau echivalent nuclear.
Originea nucleului, ca organit bine delimitat în celulă, constituie încă un subiect mult controversat. Una din ipotezele luate în considerare se bazează pe observaţiile efectuate pe celulele procariote cu un cromozom fixat la un situs specific în interiorul membranei plasmice. Se presupune că membrana plasmică, la nivelul mezozomului, prin invaginare ar fi generat învelişul nuclear în jurul cromozomului unic procariot.
În celulele eucariote, nucleul este un organit bine individualizat cu rol de depozitare a informaţiei genetice, de reglare şi de control a tuturor activităţilor celulare. Excepţie fac celulele anucleate (plăcuţele sanguine şi hematiile la om), care sunt incapabile de a sintetiza proteine.
În timpul mitozei, învelişul nuclear se dezagregă, fapt ce conduce la presupunerea că ar exista o reversie temporară a celulei eucariote la tipul procariot, cromozomii nemaifiind compartimentaţi.
Poziţia nucleului în celulă poate fi: centrală în celule tinere, sferice;excentrică / paracentrală în celulele diferenţiate (adipocite) şi deplasată la polul bazal în celulele secretoare.
Numărul nucleilor într-o celulă poate varia de la un singur nucleu în majoritatea celulelor, la 2 nuclei în hepatocite, condrocite, la 100 de nuclei în osteoclaste şi câteva sute de nuclei (fibra musculară striată).
Forma nucleului, în general, corespunde formei celulei căreia îi aparţine, putând fi:sferică, ovoidă, fusiformă sau de bastonaş, turtită, multilobată ş.a.
Dimensiunile nucleului variază în funcţie de ţesutul/ organul de referinţă, de vârsta celulei şi activitatea ei metabolică. În general, dimensiunile lui sunt de circa 5-12 gm (4 gm la spermatozoid, 20-25 gm la ovocit).
Între volumul nucleului şi cel al citoplasmei există un raport, numit raport nucleo-citoplasmatic, care poate varia între 1:3 până la 1:20. Celulele tinere cu activitate metabolică intensă au nuclei mari, cu volum mai mare faţă de cel al citoplasmei. După acest criteriu se poate stabili vârsta celulelor, dar şi diagnosticul de celule tumorale. Acestea din urmă sunt considerate celule foarte tinere, au nuclei cu morfologie variată, uneori chiar cu aspect monstruos.
Studiul nucleului în celula vie
Examinat la microscopul cu contrast de fază sau la cel cu contrast interferenţial Nomarski, nucleul are aspectul unei mase sferice, delimitată de un înveliş, la interior având un corpuscul refringent, numit nucleol.
Se ştie că la o celulă vie dacă i se perforează învelişul nuclear prin microdisecţie, micro-puncţie sau accidental, acesta nu se mai reface, iar conţinutul nucleului se amestecă cu citoplasma provocând moartea celulei respective.
Prin microfilmare s-au pus în evidenţă o serie de mişcări ale nucleului în celulă astfel:
- mişcări de deplasare datorate curenţilor citoplasmatici sau a membranelor
- ondulante,
- mişcări de rotaţie, dependente de mişcările citoplasmei,
- mişcări de expulzare din celulă, în cazul nucleilor degeneraţi.
În celula vie nucleul prezintă proprietăţi fizico-chimice specifice:
- nucleul este mai dens decât citoplasma, cu importanţă în ordinea de sedimentare a componentelor celulare în timpul ultracentrifugării diferenţiale;
- nucleul are o vâscozitate mai redusă faţă de citoplasmă;
- nucleul are un pH alcalin de 7,6.
Structura nucleului celular
Studiind nucleului fixat la microscopul optic şi la cel electronic pe secţiuni (preparate microscopice) se disting patru componente majore ale nucleului celular: învelişul nuclear, cromatina, nucleolulşi nucleoplasma.
Nucleul este delimitat de envelopa nucleară, o membrană dublă care separă materialul genetic de citoplasmă. Alte componente esențiale sunt:
- Nucleoplasma – un fluid în care se află cromatina și nucleolii.
- Cromatina – ADN-ul asociat cu proteine histonice, care se compactează în cromozomi în timpul diviziunii celulare.
- Nucleolul – o structură implicată în sinteza ribozomilor.
- Pori nucleari – canale prin care moleculele esențiale trec între nucleu și citoplasmă.
Învelişul nuclear al nucleului celular
Învelişul nuclear are o structură specifică, fiind alcătuit din două membrane concentrice, ca nişte foiţe, aflate în continuitate una cu cealaltă şi din loc în loc cu membrana reticulului endoplasmatic granular.
Pe suprafaţa membranei externe a învelişului nuclear se găsesc ribozomi ataşaţi.
Membrana internă este lipsită de ribozomi fiind în contact cu nucleoplasma. Pe faţa internă a învelişului nuclear, în zona neocupată de pori, se află o reţea proteică fibroasă, numită lamina nucleară, ce determină forma nucleului, mediază interacţiunea dintre învelişul nuclear şi cromatină, induce dispariţia şi refacerea învelişului nuclear în cursul mitozei.
Cele două membrane ale învelişului nuclear formează un sistem de membrană dublă, între ele existând un spaţiu perinuclear (perimembranar) de circa 20-40 nm prin care se transportă proteinele sintetizate în ribozomii ataşaţi învelişului nuclear.

Figura 1 - Schema nucleului interfazic (după Bloom et al., 1975)
Nucleul este un centru metabolic activ cu funcţii vitale pentru celulă prin care o serie de componente biochimice cu dimensiuni variabile trebuie să traverseze învelişul nuclear (tipuri de ARN, histone, ADN-polimeraze şi ARN- polimeraze etc.) dinspre nucleu în citoplasmă sau invers.
Traversarea învelişului nuclear este un proces selectiv efectuat prin structuri tipice, numite pori nucleari, care la mamifere apar pe circa 10% din suprafaţa învelişului nuclear, în locurile unde membrana internă se continuă cu cea externă (aproximativ 11 pori/pm ).

Figura 2 - Schema porului nuclear (după Karp, 1984)
Complexul porului se compune din porul propriu-zis, de formă octogonală, cu diametrul de circa 60-90 nm, înconjurat de un inel (annulus) format din opt subunităţi proteice (granule) de 15 nm. În centrul porului nuclear apare un canal „apos” prin care moleculele solubile pot circula în ambele sensuri.
Nucleul exportă în citoplasmă precursori ribozomali şi particule ribonucleoproteice, formate din complexe ale ARNm, ARNt şi proteine speciale-receptori, ce sunt tranzitate prin canalul porului, precum şi factori de transcripţie ce se reîntorc în citosol.
Cromatina nucleară
În nucleul celulelor eucariote, ADN-ul există sub formă de cromatină organizată în structuri mai complexe. Cromatina mai conţine mici cantităţi de ARN şi proteine nehistonice, astfel că ea poate fi considerată chimic o nucleoproteină în care proteinele histonice şi nehistonice interacţionează între ele şi cu ADN-ul.
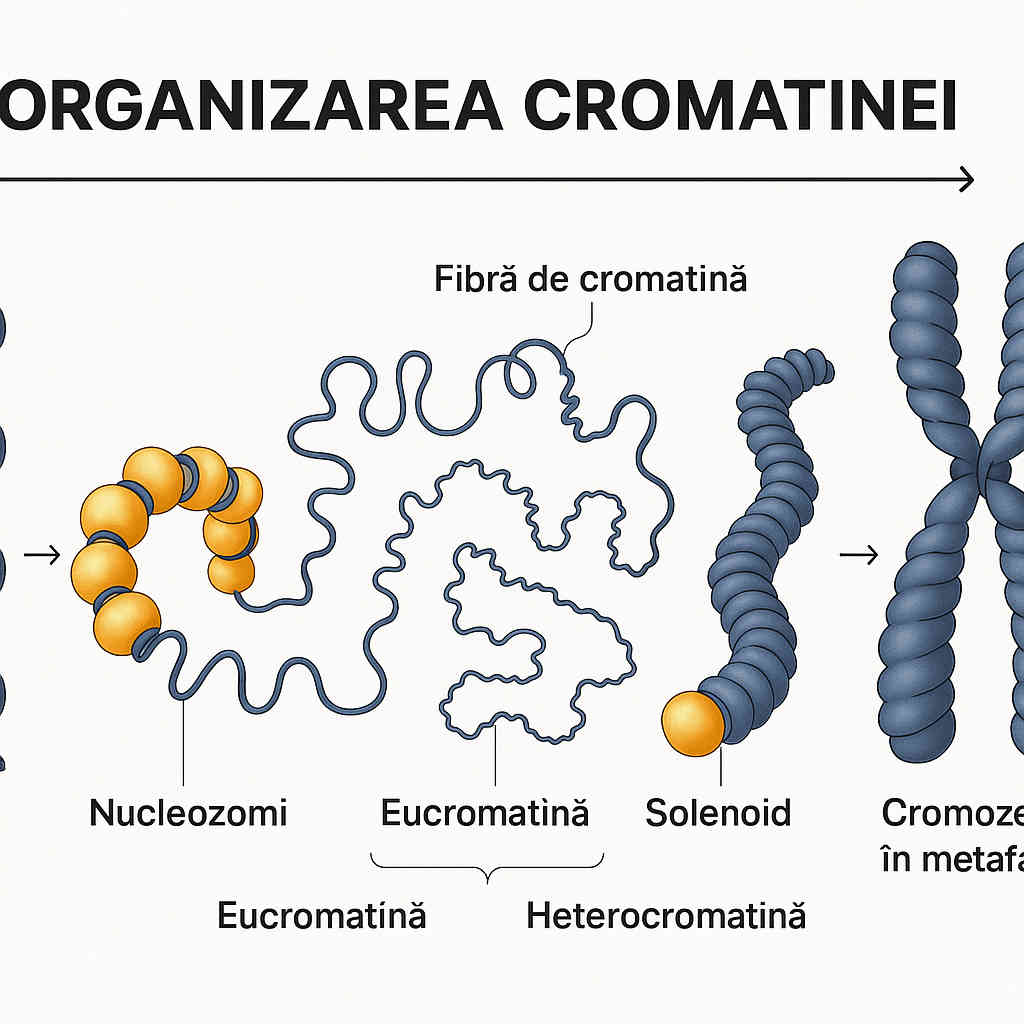
În interfază, cromatina nucleară are un aspect relaxat, adică o formă desfăşurată a cromozomilor, ei fiind astfel invizibili la microscopul optic obişnuit. Cromatina celulei eucariote are la bază nucleozomii [Arthur Kornberg 1973], fiecare fiind constituit dintr-un complex de 8 molecule de histone (două copii a 4 tipuri de histone: H2A, H2b, H3 şi H4 ), înconjurat de un segment de ADN format din circa 147 pb (146-166 perechi de baze).
Celulele umane conţin aproximativ 3x107 nucleozomi, ei reprezentând primul nivel ierarhic de organizare a cromatinei. De la nivelul a 10-90 pb de ADN un nucleozom se leagă de cel următor, fiecare regiune de legare fiind ocupată de o singură moleculă de histonă Hi.
Nucleozomii sunt legaţi între ei în „şirag de perle” în structuri mult mai compacte, spiralizate, cu diametrul de 250-300 Â, numite fibre groase. Acestea conţin circa 45.000 pb şi reprezintă al doilea nivel de condensare al cromatinei. Prin studii fizico-chimice s-a sugerat că aceste fibre ar consta dintr-o împachetare continuă a nucleozomilor pentru a forma un solenoid.
Fiecare solenoid este un helix de 6 până la 9 nucleozomi ce conţin circa 1.200 pb de ADN, structură în care gradul de împachetare creşte foarte mult pentru a se realiza starea de condensare a cromozomilor din metafază.

Figura 3 - Componentele nucleozomului (după Mescher, 2010)
În metafază, când se formează cromozomii, fibrele groase se condensează mai mult, iar prin împachetări adiţionale apar structuri mai complexe, numite domenii (loops) cu 35.000-90.000 pb de ADN. La om cele 46 de molecule de ADN, totalizând peste 1 metru în lungime, necesită plieri şi compactări multiple pentru a încăpea într-un nucleu cu diametrul de circa 10 pm.
Aceste procese pot avea loc datorită interacţiunilor dintre histone (cozile lor), ce determină nucleozomii să se asocieze în fibre compacte, care se pliază în structuri mai complexe cu configuraţii incerte, modificabile probabil în funcţie de nivelul de activitate al genelor din zonă. Acest tip structural de cromatină este bine legat într-un suport de organizare nucleară, numit matrix nuclear.
În interfază, cromatina nucleară are un aspect relaxat, adică o formă desfăşurată a cromozomilor, ei fiind astfel invizibili la microscopul optic obişnuit.
Evidenţieri prin colorare
Cromatina se colorează intens cu coloranţi bazici (ex, hematoxilină, albastru de tripan) datorită conţinutului mare de ADN. Aspectul microscopic al cromatinei este variat putând fi de grămezi neregulate, granule sau de reţele de filamente mai mult sau mai puţin fine, care în anumite zone (puncte de intersecţie) sunt mai groase.
În interfază, cromatina prezintă două aspecte tinctoriale [după Emil Heitz, 1929](fig. 1), astfel:
Eucromatina are aspect de reţea de filamente foarte fine (30 nm grosime), slab colorabile, este metabolic activă conţinând genele responsabile de transcripţia ARN. Filamentele de eucromatină nu au un diametru constant, prezentând pe traiectul lor umflături ce corespund zonelor în care cromatina este mai des înfăşurată.
Eucromatina se clasifică în:
- eucromatină activă , adică fracţiunea de eucromatină pe care se desfăşoară continuu procesul de transcripţie şi care asigură viaţa normală a celulei;
- eucromatina permisivă , adică fracţiunea de eucromatină potenţial activă, care în urma unor semnale specifice modulatoare (ex., hormoni) devine activă.
Heterocromatina apare sub formă condensată şi colorată intens, uneori ia aspect de corpusculi numiţi cromocentrii sau cariozomi, fiind inactivă metabolic. Inactivitatea metabolică a fost demonstrată prin administrarea în celule a unui precursor de ARN marcat radioactiv, numit [3H] uridină.
Celulele respective au fost apoi fixate, secţionate şi autoradiografiate observându-se că heterocromatina lor nu a fost specific marcată, indicând absenţa capacităţii de sinteză de ARN.
Secţiunile heterocromatice ale cromozomului pot fi de două tipuri, cu heterocromatină constitutivă şi cu heterocromatină facultativă, în funcţie de condensarea permanentă a cromatinei sau de condensarea ei numai în anumite condiţii. ADN-ul heterocromatinei constitutive este definitiv inactivat şi rămâne în stare condensată tot timpul, astfel că aceleaşi regiuni de pe cromozomii omologi vor fi inactive definitiv.
Heterocromatina facultativă este o cromatină condensată ce conţine gene structurale represate (inactive), pe care nu se desfăşoară procesul de transcripţie deşi într-o perioadă anterioară s-a făcut sau nu transcripţie, iar dacă se transformă temporar în eucromatină (sub influenţa unui „depresor”) se va face sau nu transcripţia.
În funcţie de tipul de cromozomi căruia îi corespunde, heterocromatina facultativă poate fi:
- heterocromatina facultativă autosomală corespunde porţiunilor condensate heterocromatice din perechile de cromozomi autosomi. Aceste porţiuni conţin lanţuri de gene structurale inactive transcripţional. Aceste gene structurale devin active, dar nu toate deodată, ci numai în anumite condiţii ale dezvoltării şi diferenţierii celulare (ex., în celulele precursoare pentru hematii, în transformarea limfoblastică a limfocitelor).
- heterocromatina facultativă gonozomală (corpuscul Barr, cromatina de sex, cromatina X) este un corpuscul cromatinian (cromocentru) prezent în mod normal numai în nucleii celulelor somatice ale indivizilor de sex femel.
Cromozomul X este implicat în determinarea sexului şi a fost descris pentru prima oară în 1949 de către Murray L. Barr şi Ewart G. Bertram ca „marker citologic pentru recunoaşterea sexului”. Corpusculul Barr măsoară circa 1 pm, are formă plan-convexă, convex-concavă, biconvexă sau triunghiulară, iar în funcţie de tipul celular poate fi localizat în nucleu, în apropierea membranei interne a acestuia (în celulele mucoasei bucale, celulele epidermului) sau ataşat nucleolului (în celulele nervoase).
Dimorfismul sexual heterocromatinian este foarte evident în unele leucocite, numite granulocite neutrofile, prin prezenţa corpusculului Barr cu aspect de băţ de tobă (drum stick, nodul de sex), ataşat printr-un filament subţire de unul dintre lobii nucleului.La indivizii masculi umani, nodulii de sex sunt absenţi sau foarte rari, iar la femeile normale frecvenţa lor este de 7/500 de granulocite neutrofile sau chiar mai mult.

Figura 4 - Aspectul cromatinei sexuale în epiteliul bucal si într-un leucocit polimorfonuclear (după Mescher, 2010)
Corpusculul F (cromatina Y) poate fi observat în lumină ultravioletă după colorarea cu fluorocromi a cromozomilor (pentru identificare prin procedeul de bandare) în nucleii celulelor de la bărbaţi normali. A fost descoperit de Pearson în anul 1970 şi constituie expresia citologică a cromozomului Y, adică a porţiunii distale a braţelor lungi ale cromozomului. La indivizii cu doi cromozomi Y se observă doi corpusculi F, în aceste cazuri fiind posibilă detectarea rapidă a indivizilor XYY, care statistic par predispuşi unor comportamente antisociale periculoase. Cu ajutorul acestei metode pot fi identificaţi şi spermatozoizii Y.
Cromatina celulelor eucariote mai conţine proteine nehistonice, proteine reglatoare, factori de transcripţie, enzime de tip ADN-polimeraza şi ARN- polimeraza, enzime de reparare, topoizomeraze ş.a.
Nucleolul (nucleolus lat.)
Nucleolul este prezent în nucleul tuturor celulelor eucariote, cu excepţia celulelor embrionare din care lipseşte, nefiind iniţiată sinteza proprie de proteine. Nucleolul a fost observat pentru prima dată de Felice Fontana în 1781, în anumite celule epidermice, în nucleul cărora apărea ca „o pată mai densă". În anul 1836, Gabriel Valentin a descris nucleolul ca fiind „o granulă densă în interiorul nucleului"".
Spre sfârşitul secolului 19 s-a făcut asocierea între prezenţa nucleolului şi activitatea de sinteză a celulei, când s-a observat că nucleolul este voluminos în celulele secretorii şi în ovocite sau este mic, chiar absent în celulele musculare sau spermatozoizi.
După anul 1960 s-a precizat că nucleolul joacă un rol esenţial în biogeneza ribozomilor, fiind sediul unui considerabil trafic molecular intracelular, adică de transcripţie a ARNr, şi de asamblare a subunităţilor ribozomale.
Nucleolul este o structură intranucleară fără membrană proprie, cu dimensiuni de 1-2 pm, de formă sferică sau ovoidală, fiind considerată cea mai densă structură din celulă (densitate 1,35). În nucleu, nucleolii pot fi poziţionaţi central sau paracentral, în raport cu starea funcţională a celulei. În celulele fixate şi colorate cu hematoxilină-eozină, nucleolul apare ca un corpuscul bazofil, mai ales în nucleii eucromatici.

Figura 5 - Structura nucleolului (după Diculescu et al., 1983)
La microscopul electronic s-au observat trei componente nucleolare (mai rar patru), astfel:
- componenta fibrilară (pars fibrosa, filamentosa) cuprinde filamente subţiri de 55 nm grosime şi 30-40 nm lungime, grupate în pachete şi organizate ca o reţea, reprezintă situs-urile de transcripţie activă a ARN-ului de pe genele ARNr; ’
- componenta granulară (pars granulosa) este alcătuită din particule sau granule cu diametru de circa 15-20 nm, considerate precursori ribozomali, reprezintă situs-urile de asamblare a ARNr, ARN-5S şi a proteinelor ribozomale în subunităţi ribozomale;
- componenta cromozomală (pars chromosoma) este alcătuită din filamente de 10 nm grosime, repartizată la periferia nucleolului (cromatina perinucleolară) şi pătrunde spre interiorul lui sub formă de benzi. Pe unele imagini de microscopie electronică această componentă pare să dea contur nucleolului, delimitându-l de reţeaua înconjurătoare de cromatină;
- componenta amorfă (pars amorpha) este nestructurată, omogenă şi de densitate medie la fluxul de electroni. În prezent se mai discută dacă este o componentă reală a nucleolului sau este carioplasma ce umple spaţiile libere.
Compoziţia chimică a nucleolului constă în prezenţa de ADN (3%), ARN (7%) şi proteine (90%) în proporţii uşor variabile din greutatea uscată, în raport de tipul celular şi de momentul funcţional ales, complexe ARN-proteine, cantităţi infime de minerale (Ca, Mg, Zn, Co etc.) şi urme de lipide.
ARN-ul nucleolar este în cantitate relativ mare, deoarece în nucleol se sintetizează (transcrie) ARNr, iar pentru ARNm şi ARNt nucleolul este o staţie intermediară obligatorie în tranzitul lor spre citoplasmă.
Raportul nucleol-nucleu
În general, nucleolul reprezintă cam 3% din volumul nucleului, iar cu cât celula este mai activă în sinteza de proteine cu atât acest raport va avea o valoare mai mare. Este considerat un criteriu important în stabilirea vârstei celulei şi în confirmarea stării de malignitate. În celula tânără nucleul este mare, eucromatic, cu 1-2 nucleoli evidenţi şi citoplasmă puţină.
Biogeneza nucleolilor
Nucleolul parcurge un ciclu propriu, fiind vizibil în interfază, dispare în profază şi reapare în telofază. Există o concepţie a ’’ organizatorilor nucleolari” conform căreia ei ar fi într-o strânsă relaţie topografică cu nucleolii.
Există o serie de ipoteze clasice privind biogeneza nucleolului:
- ipoteza continuităţii , conform căreia nucleolul ar persista în cursul mitozei sub formă de granule şi/sau fibrile subţiri (pseudonucleoli, corpusculi prenucleolari), ataşate de cromozomi, împreună cu care se vor repartiza în celulele-fiice, fiind asamblate de organizatorii nucleolari;
- ipoteza formării de novo , conform căreia materialul nucleolar se dezintegrează complet în timpul mitozei, nefiind încorporat în noii nucleoli.
Concepţia actuală asupra biogenezei nucleolului afirmă că ADNr (componenta cromozomală, organizatorul nucleolar) trebuie să se transmită de la o generaţie celulară la alta, în timp ce celelalte componente moleculare (fibrilară, granulară, amorfă) nu necesită perpetuarea pentru că pot fi oricând sintetizate pe baza informaţiei conţinute în ADN, în faza Gi de ciclu celular când cistronii sunt reactivaţi pentru transcripţie.
Funcţiile nucleolului
Nucleolul are rol în sinteza de ARNr şiîn biogeneza ribozomilor; întransferul de ARNm şi ARNt spre citoplasmă şi în pregătirea mitozei.
Matricea nucleară (carioplasma, sucul nuclear)
Matricea nucleară constituie scheletul de natură proteică ce înglobează cromatina şi nucleolii şi care se sprijină la exterior pe foiţa internă a învelişului nuclear. Matricea nucleară este o reţea fibroasă ce conţine aproximativ 10% din totalul proteinelor nucleare, 30% din ARN-ul nuclear, 1-3% din ADN-ul total şi 3% din fosfolipidele nucleare.
Această structură a fost pusă în evidenţă în anul 1987 de către Peter Lawrence, după ce nucleii au fost trataţi cu nucleaze pentru îndepărtarea acizilor nucleici şi cu detergenţi şi săruri pentru îndepărtarea proteinelor solubile. În final, a rămas o reţea de proteine filamentoase interconectate, adică matricea nucleară.
La microscopul electronic, matricea nucleară apare ca o reţea pe toată aria nucleului, formată din particule electrono-dense de circa 15-20 nm în diametru, numite particule sau granule matriceale şi pachete dense de fibrile, cu diametrul fibrilelor de circa 5 nm, numite fibrile matriceale. Această reţea matriceală se află în relaţii de continuitate cu lamina densa interna şi cu complexul porului aflate pe faţa nucleară a membranei interne a învelişului nuclear.
Matricea nucleară are rol esenţial înorganizarea nucleului, în sinteza de ADN şi ARN, înmedierea unor semnale hormonale, rol de suport pentru depozitarea granulelor ribozomale ş.a.
Cromozomii (chroma gr.= culoare, soma gr.= corp)
Cromozomii au fost descrişi pentru prima dată în 1848 de către Wilhem Hoffmeister la plante, iar în 1882 Walther Flemming a observat că în cursul diviziunii celulare cromatina se condensează sub formă de bastonaşe, ce clivează apoi în sens longitudinal.
Denumirea de cromozomi a fost dată de către Heinrich Wilhelm von Waldeyer-Hartz în anul 1888.
Abia în secolul 20 s-a demonstrat rolul important al cromozomilor în transmiterea ereditară a caracterelor şi structura intimă a moleculei de ADN (1910, Thomas Hunt Morgan; 1953, James Watson şi Francis Crick).
Cromozomii conţin majoritatea materialului genetic dintr-o celulă, fiind destinaţi pentru stocarea, transmiterea, exprimarea şi evoluţia informaţiei genetice proprii.
În interfază, cromozomii nu sunt vizibili la microscopul optic obişnuit, deoarece cromatina este dispersată în toată nucleoplasma.
Începutul diviziunii celulare se manifestă prin apariţia cromozomilor sub formă de filamente subţiri (bastonaşe) în interiorul nucleului, cu lungimi de 0,2-5 pm şi diametru de 0,2-2 pm. În timpul profazei şi metafazei cromozomii sunt formaţi din câte două filamente identice înfăşurate, numite cromatide, fiecare din ele conţinând una sau două molecule-fiice identice de ADN.
Cromatidele devin mai condensate datorită filamentelor nucleoproteice spiralate, numite cromoneme ce sunt variabile numeric în funcţie de lungimea cromozomului.
Cromatidele-surori sunt menţinute unite la nivelul constricţiei printr-o structură specifică numită centromer [Heinrich Wilhelm von Waldeyer-Hartz, 1903]. S-a stabilit că fiecare centromer are doi kinetocori, câte unul pentru fiecare cromatidă, ca loc de ataşare pentru microtubulii fusului de diviziune [Friedrich Scharader 1936].
Extremitatea terminală a cromatidei este rotunjită, poartă denumirea de telomer [Hermann Muller, 1932], are proprietăţi genetice specifice, prevenind fuzionarea cromozomilor (le conferă individualitate).
În ultimii ani s-a discutat tot mai mult de implicarea telomerului în durata de viată a celulei, adică de numărul de diviziuni pe care le poate parcurge o celulă în condiţii normale [Leonard Hayflick]. Se pare că în celulele canceroase se activează o enzimă, numită telomerază, care adaugă o cantitate de ADN la telomer cu fiecare diviziune, fapt ce ar explica mitozele succesive, fără control.

Figura 6 - Model de cromozom metafazic (după Voiculeţ and Puiu, 1997)

Figura 7 - Cromozom eucariot (după Becker et al., 2009)
În funcţie de poziţia centromerului, cromozomii monocentrici (cu un singur centromer) pot fi de 4 feluri: metacentrici, submetacentrici, acrocentrici şi telocentrici.
Numărul cromozomilor variază în funcţie de specie, tip celular, condiţii de apariţie a unor mutaţii genetice ş.a.
Tabel 1 Numărul cromozomilor în celule somatice (date din Strickberger, 1976; Darnell et al., 1988)
| Specia considerată | 2n | Specia considerată | 2n |
| Om (Homo sapiens) | 46 | Câinele (Canis familiaris) | 78 |
| Maimuţa Rhesus (Macaca mulata) | 42 | Găina (Gallus domesticus) | 78 ± |
| Gorila (Gorilla gorilla) | 48 | Calul (Equus calibus) | 64/67 |
| Vaca (Bos taurus) | 60 | Broasca (Rana pipiens) | 26 |
| Capra (Capra prisca) | 60 | Vierme de mătase (Bombyx mori) | 56 |
| Oaia (Ovis aries) | 54 | Crapul (Cyprinus carpio) | 104 ± |
| Iepure de casă | 44 | Musculiţa de oţet | 8 |
| (Oryctolagus cuniculus) | (Drosophila melanogaster.) | ||
| Porcul (Sus scrofa domesticus) | 40 | Albina domestică (Apis melifera) | 32 |
| Şoarecele (Mus musculus) | 40 | Furnică (Myrmecia pilosula) | 2 |
| Pisica (Felis domesticus) | 38 | Algă verde(Acetabularia mediterranea) | 20 ± |
| Şobolanul (Rattus norvegicus) | 42 | Porumb (Zea mays) | 20 |
| Hamsterul (Mesocricetus auratus) | 44 | Cartof (Solanum tuberosum) | 48 |
| Cobaiul (Cavia cobaya) | 64 | Fluturi (diverse specii) | ~380 |
Cariotipul reprezintă totalitatea caracterelor morfologice dintr-un set cromozomal, caracteristic pentru fiecare specie în parte [Cyril D. Darlington, 1939], iar cunoaşterea cariotipului normal pe specii ajută la identificarea unor boli ereditare sau dobândite în urma unor mutaţii ce implică numărul sau structura cromozomilor (translocaţii).
Incluziunile nucleare
În situaţii foarte rare, accidentale sau de boală, nucleul celular poate conţine substanţe ce nu intră în compoziţia sa obişnuită, cum ar fi: particule de citoplasmă cu resturi de mitocondrii sau de reticul endoplasmatic, substanţe chimice (ex., glicogen, lipide, granule de secreţie), diverşi corpi cu aspect cristalin (cristaloizi).
La plante, s-a semnalat destul de des prezenţa unor cristaloizi cu diferite forme de bastonaşe, sferici, ovali sau cu margini neregulate. La animale s-au observat cristaloizi în celulele ganglionare ale lui Herisson sau în intestinul unor nevertebrate (gândacul-morar). În anumite condiţii s-a observat că hemoglobina cristalizează în nucleii celulelor hepatice, arătând ca o albumină străină de structura nucleului celular.
Se pare că în anumite condiţii de activitate celulară (sinteză, secreţie) o serie de substanţe accesorii pot fi mai abundente, cu aspecte şi forme mai mult sau mai puţin definite, asemănătoare fie nucleolilor, fie cristaloizilor.
Într-o stare de suferinţă (boală) ele pot avea semnificaţie de diagnostic:
- prezenţa lipidelor sub formă de picături indică intoxicaţii severe,
- prezenţa incluziunilor virale, ca formaţiuni cristaloide, indică o stare de degenerescenţă precoce a celulei.
Funcțiile nucleului celular
Nucleul joacă un rol esențial în menținerea și transmiterea informației genetice. Principalele sale funcții includ:
a) Stocarea și protejarea ADN-ului
Nucleul păstrează ADN-ul, protejându-l de degradarea enzimatică și reglementând accesul la informația genetică.
b) Controlul activității celulare
ADN-ul conține instrucțiuni pentru sinteza proteinelor, care sunt transmise prin ARN mesager (ARNm). Acest proces controlează metabolismul și dezvoltarea celulei.
c) Reglarea expresiei genelor
Nucleul decide când și cum sunt activate sau reprimate anumite gene, influențând funcțiile și caracteristicile celulare.
d) Sinteza ribozomilor
Nucleolul produce ARN ribozomal (ARNr), esențial pentru formarea ribozomilor, structurile care sintetizează proteinele.
e) Diviziunea celulară
În timpul mitozei și meiozei, nucleul coordonează replicarea ADN-ului și distribuirea corectă a materialului genetic către celulele fiice.
Importanța nucleului celular
Nucleul este vital pentru supraviețuirea celulei și pentru buna funcționare a organismului. Defectele nucleare pot duce la mutații genetice, cancer sau boli genetice.
În concluzie, nucleul celular este un organit esențial pentru celulele eucariote, având rolul de a proteja ADN-ul, a coordona metabolismul și a regla diviziunea celulară. Studierea sa este fundamentală în domeniul biologiei moleculare și al medicinei genetice.
Întrebări frecvente despre nucleul celular
1. Ce este nucleul celular și ce rol are în celulă?
Nucleul celular este un organit membranos prezent în celulele eucariote, care stochează materialul genetic (ADN) și controlează procesele celulare esențiale, precum sinteza proteinelor și diviziunea celulară.
2. Care sunt componentele principale ale nucleului celular?
Componentele principale sunt: învelișul nuclear, cromatina, nucleolul, nucleoplasma și porii nucleari. Acestea colaborează pentru protejarea și utilizarea eficientă a ADN-ului.
3. Ce este cromatina și cum se diferențiază eucromatina de heterocromatină?
Cromatina este o structură formată din ADN și proteine histonice. Eucromatina este slab condensată și activă transcripțional, în timp ce heterocromatina este densă și inactivă metabolic.
4. Ce funcții îndeplinește nucleolul în cadrul nucleului?
Nucleolul este responsabil de sinteza ARN-ului ribozomal (ARNr) și asamblarea subunităților ribozomale, esențiale pentru producerea proteinelor.
5. De ce este importantă poziția și dimensiunea nucleului într-o celulă?
Poziția și dimensiunea nucleului reflectă vârsta și activitatea metabolică a celulei. Un nucleu mare și eucromatic indică o celulă activă, în timp ce anomaliile pot semnala afecțiuni precum cancerul.